 Вход
ВходДекоративно-растущие многолетние растения – семейство губоцветные. Порядок губоцветные (Lamiales) (Н. Н. Цвелев)
-Представители этого семейства легко узнаются уже по строению венчика цветков, имеющего длинную трубку и двугубый зев, напоминающий разинутую пасть сказочного животного. Хотя сходный по облику венчик имеется еще в нескольких семействах цветковых растений (например, у многих норичниковых), для губоцветных он особенно характерен. Не менее своеобразен и плод губоцветных, состоящий из 4 односемянных орешкообразных или редко костянковидных долей, тогда как у сходных с ними по строению венчика норичниковых плод - обычно многосемянная коробочка. К числу важных отличительных особенностей губоцветных принадлежат также супротивные (очень редко мутовчатые), обычно цельные листья без прилистников и обычно четырехгранные стебли. Очень существен ароматический запах, свойственный большинству видов губоцветных, который определяется присутствием на всех или на некоторых частях растения желёзок, выделяющих эфирные масла сложного состава (в них входят ароматические спирты, фенолы, терпены, альдегиды и другие органические соединения). Именно присутствием этих масел в значительной степени определяется практическое использование губоцветных в качестве технических, лекарственных и ароматических растений.
, 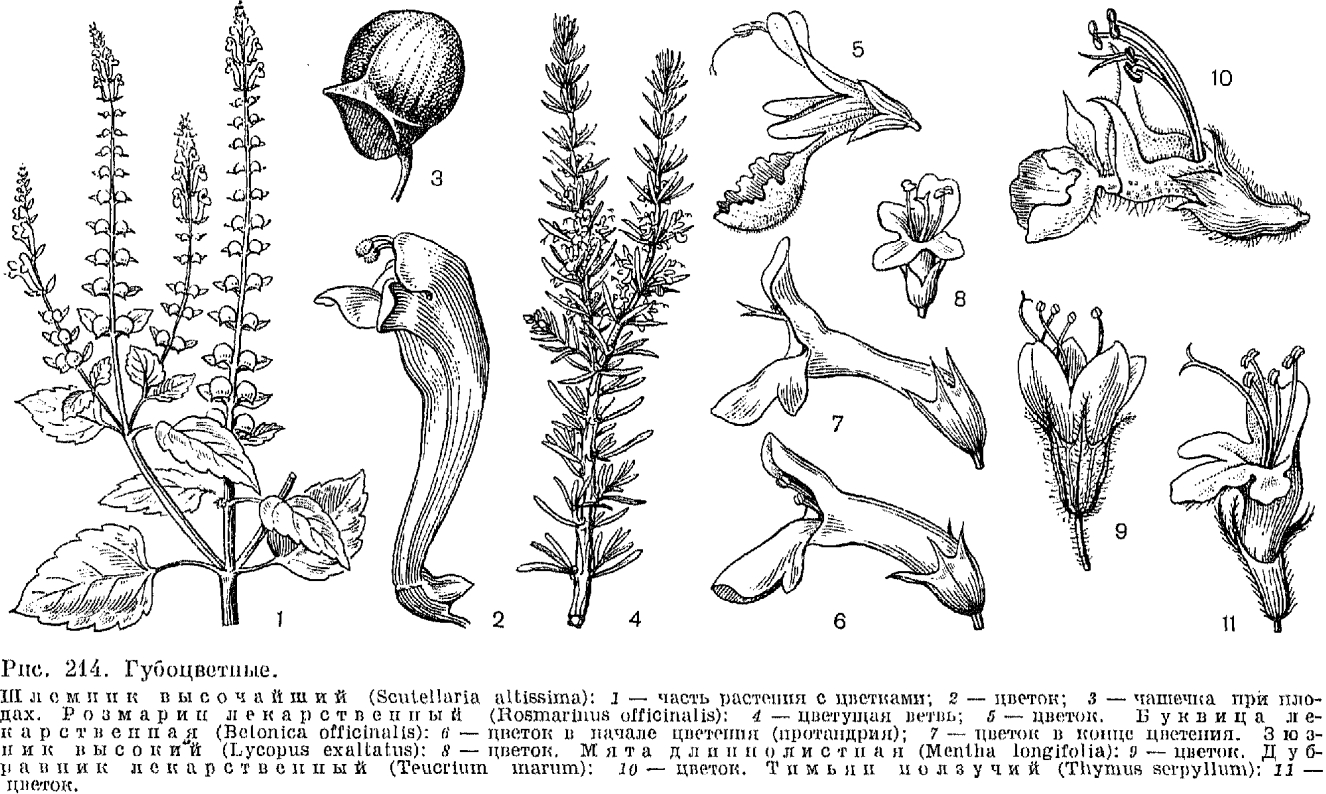
Большинство губоцветных - травы и полукустарники. Однако среди них, особенно в тропиках и субтропиках, много и кустарников, примером которых может служить распространенный в Средиземноморской флористической области розмарин лекарственный (Rosmarinus officinalis, табл. 55) - вечнозелёный кустарник с мелкими линейными листьями и сине-фиолетовыми (до почти белых) цветками (рис. 214). Губоцветные - деревья имеются только в тропиках, но, в отличие от близкого преимущественно древесного семейства вербеновых, ими являются лишь немногие виды двух американских родов: хиптис (Hyptis) и левкосцептрум (Leucosceptrum). «Чемпион» по высоте среди них - бразильский вид хиптис перепончатый (H. membranacea), достигающий в высоту 12-15 м, в то время как другие древесные губоцветные обычно не достигают высоты 5 м. В тропиках встречаются и немногие лианы, к которым принадлежат лишь американский род салазария (Salazaria), некоторые виды шлемника (Scutellaria, табл. 55) и гавайского рода стеногина (Stenogyne).
Стебли травянистых губоцветных обычно прямостоячие и не нуждаются в опоре, хотя имеются виды со стелющимися по земле и укореняющимися в узлах стеблями (например, будра плющелистная - Glechoma hederacea). У живучки ползучей (Ajuga reptans), кроме прямостоячих репродуктивных побегов, из пазух листьев розетки образуются дугообразные направленные к земле и укореняющиеся верхушками вегетативные побеги, подобные усам земляники. Хорошо развитая розетка прикорневых листьев, сохраняющаяся во время цветения растения, имеется у многих травянистых губоцветных (например, у некоторых шалфеев - Salvia).
Главный корень часто сохраняется в течение всей жизни растения, реже отмирает и замещается придаточными корнями, отходящими или от основания стебля или от отходящих от него ползучих подземных побегов - корневищ, свойственных многим видам губоцветных. Довольно редко среди губоцветных встречаются виды с корневыми отпрысками, например живучка женевская (Ajuga gennevensis, табл. 55). У многих прибрежных видов, обитающих на переувлажненных местообитаниях, в корневищах образуются воздухоносные полости или участки воздухоносной ткани. Некоторые губоцветные имеют клубневидно утолщенные корни, в тропических странах употребляемые в пищу.
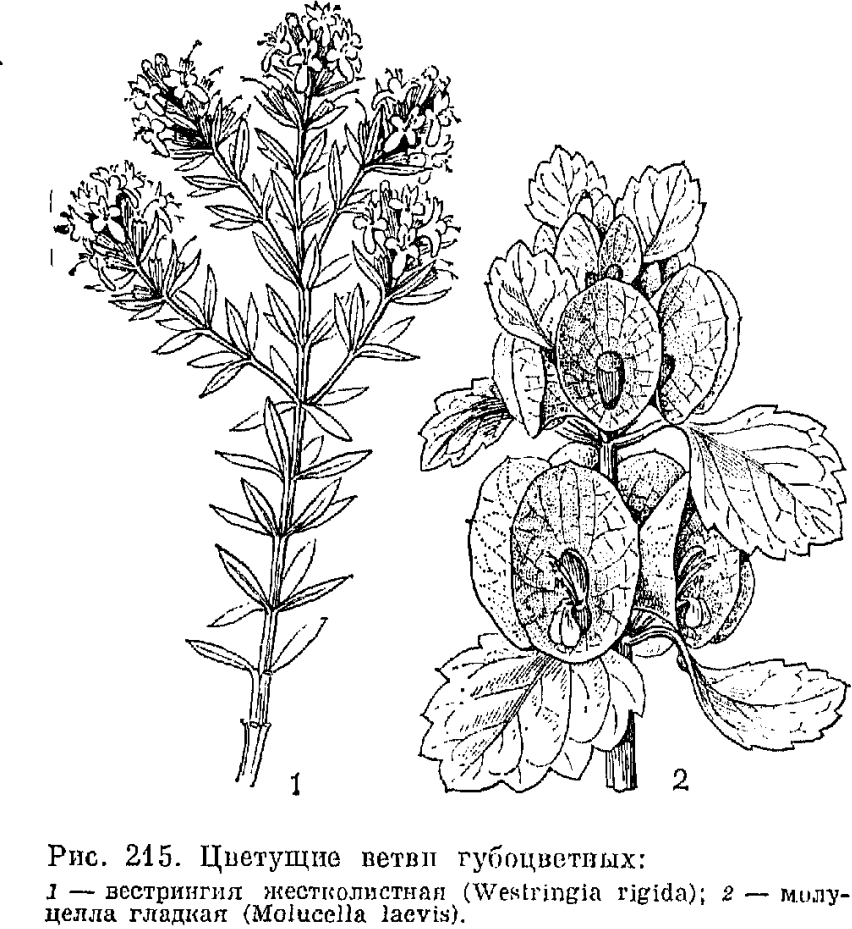
При супротивном расположении листьев близлежащие пары крестообразно чередуются друг с другом. Следствием этого и является четырехгранность стеблей губоцветных, причем грани могут быть плоскими, выпуклыми или вогнутыми. Видов, имеющих мутовчатое расположение листьев, среди губоцветных немного. К ним принадлежит, в частности, оригинальный австралийский кустарник вестрингия (Westringia) с мелкими цельнокрайними листьями, расположенными мутовками по 3-6 (рис. 215). Очередное листорасположение отмечено только у первых листьев проростков в родах зопник (Phlomis) и буквица (Betonica).
Листья губоцветных обычно цельные и часто цельнокрайние, хотя встречаются и перистораздельные (например, у шалфея скабиозолистного - Salvia scabiosifolia). Известны как голые или почти голые виды, например декоративный шалфей блестящий (S. splendens), так и виды, густо покрытые волосками. Среди последних такие средиземноморские виды, как чистец критский (Stachys cretica) и железница крымская (Sideritis taurica), не уступают по красоте знаменитому альпийскому эдельвейсу. Из волосков наиболее распространены многоклеточные простые волоски. Головчатые волоски, головка которых функционирует как желёзка, вырабатывающая эфирное масло, встречаются у многих губоцветных.
Обычно пятичленные и, как правило, обоеполые цветки губоцветных располагаются в пазухах неизмененных или видоизмененных в прицветники листьев. Лишь в немногих случаях (например, у видов шлемника) они одиночные, обычно же цветки образуют так называемые ложные мутовки, составленные двумя супротивными соцветиями, несущими прицветнички. Оси всех первичных соцветий, часто сильно укороченные и составляющие их цветки, кажутся расположенными непосредственно в листовых пазухах, образуя колосовидные соцветия. Иногда ось общего соцветия сильно укорочена и все соцветие становится головкообразньм, например у зизифоры головчатой (Ziziphora capitala) и у большого американского рода хиптис. У нередкого в горных районах Средней Азии зайцегуба (Lagochilus) расположенные при основании ложных мутовок прнцветнички видоизменены в мощные колючки, защищающие растение от поедания травоядными животными. У некоторых других губоцветных в колючки видоизменяются прицветники или верхние листья, а иногда зубцы листьев.
И чашечка, и венчик губоцветных обычно образованы 5 сросшимися своей основной частью в трубку листочками. Лишь немногие роды, например западносреднземноморский род преслия (Preslia), имеют четырехчленный околоцветник. Чашечка губоцветных может иметь самую разнообразную форму: трубчатую, колокольчатую, ворончатую, шаровидную, а в зеве может быть как двугубой без каких-либо зубцов, так и 5(4)-зубчатой с зубцами одинаковой или различной длины (в последнем случае чашечка также более или менее двугубая). В отличие от венчика, видоизменения которого связаны с приспособлением к опылению, видоизменения чашечки обычно связаны с приспособлением к распространению плодов. Нередко (например, у пустырника) зубцы чашечки имеют вид отогнутых в сторону колючек. Иногда вся чашечка или ее зубцы сильно разрастаются, увеличивая парусность чашечки при распространении семян с помощью ветра, или чашечка становится ярко окрашенной, играя роль в привлечении насекомых или птиц-опылителей, например, ярко-красная чашечка шалфея блестящего. Очень оригинальна чашечка у большого (около 300 видов), почти космополитного рода шлемник. Она здесь двугубая с цельнокрайними губами и после созревания плода распадается иа 2 части, имеющие вид створок: нижнюю остающуюся и верхнюю опадающую. Верхняя часть чашечки у многих видов этого рода несет еще поперечную чешуевидную складку - щиток. Шлемник имеет еще ряд особенностей, отличающих этот род от других родов губоцветных (в том числе отсутствие эфиромасличных желёзок), и не случайно некоторые авторы даже предлагали выделить его в особое семейство шлемниковые (Scutellariaceae).
,
Как уже упоминалось выше, венчик губоцветных обычно разделен на две губы, из которых верхняя образована 2, а нижняя - 3 лепестками. Верхняя губа может быть плоской или выпуклой, иногда она бывает цельнокрайней, так что никаких следов присутствия 2 лепестков не обнаруживается. Нижняя губа почти всегда более крупная (посадочная площадка для опылителей), трехлопастная с более крупной и нередко, в свою очередь, двулопастной средней лопастью. Иногда на ее боковых лопастях имеются нитевидные придатки, как у яснотки (Lamium). Довольно оригинальное строение венчика у родов дубравник (Teucrium) и живучка (Ajuga, табл. 55). У первого из них верхней губы нет совсем и тычинки вместе со столбиком далеко выступают из зева венчика (рис. 214). Однако 2 верхние лопасти, обычно образующие верхнюю губу, здесь не исчезли, а присоединены к нижней губе венчика, составленной не из 3, а из 5 лопастей. У живучки верхняя губа очень короткая по сравнению с длинной нижней губой и венчик также кажется одногубым. У базилика (Ocimum) и близких родов верхняя губа венчика образована не 2, как обычно, а 4 лепестками. Нижняя губа состоит всего из одного плоского или вогнутого лепестка. Для родственного базилику шпороцветника (Plectranthus), заходящего на юг Дальнего Востока, характерно, кроме того, наличие вздутия в нижней части трубки венчика, причем у некоторых видов это вздутие переходит в настоящую шпору. Некоторые роды губоцветных, в том числе и зюзник (Lycopus, рис. 214), имеют короткий и почти актиноморфный венчик с 4-5 лопастями. Окраска венчиков у губоцветных может быть розовой, лиловой, сиреневой, синей, желтой, белой, часто в различных комбинациях.
Тычинок в цветках губоцветных обычно 4, прикрепленных к трубке венчика. У тропического рода колеус (Coleus, табл. 55) и некоторых близких родов тычиночные нити срастаются друг с другом, образуя короткую трубку. Иногда имеется рудимент пятой тычинки, вероятно, исчезнувшей в результате преобразования актиноморфного венчика в зигоморфный в ходе эволюции губоцветных. Пара задних тычинок обычно короче передней пары, но иногда, например у котовника (Nepeta), имеет место обратное соотношение. У мяты (Mentha) с ее почти актиноморфным околоцветником все 4 тычинки почти одинаковой длины. Редукция тычинок в пределах семейства идет еще дальше - до 2 тычинок, причем редуцируются 2 задние тычинки, иногда сохраняясь в виде стаминодиев. Две тычинки характерны, например, для средиземноморского рода розмарин, шалфея и североамериканско-мексиканского рода монарда (Monarda). Ниже места прикрепления тычинок нередко имеется волосистое кольцо - защитное приспособление для нектара.
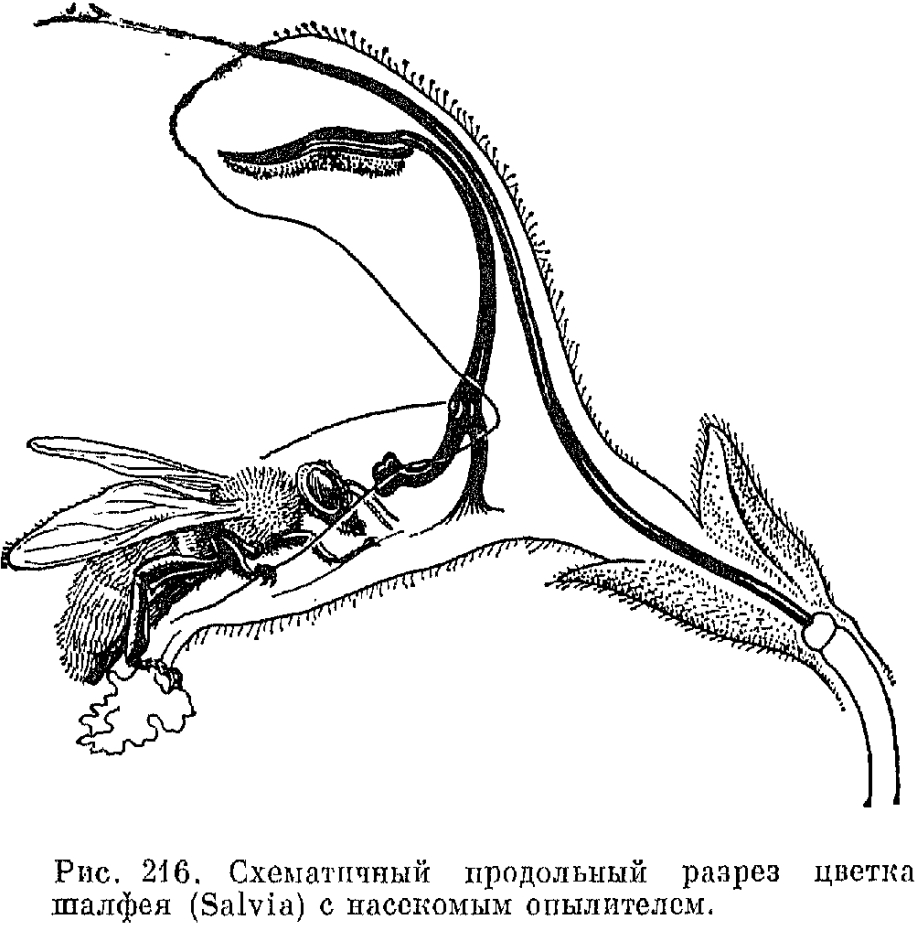
Пыльники губоцветных имеют различную форму. Гнезда их обычно одинаково развиты, реже одно из них (чаще переднее) развито слабее другого или редуцировано. У многих видов шалфея специализация тычинок зашла наиболее далеко в связи с очень совершенным приспособлением цветков к опылению определенными насекомыми (рис. 216). Каждый из пыльников двух имеющихся здесь тычинок превратился в своеобразное рычажное устройство, на одном конце которого находится вполне развитое верхнее гнездо пыльника, а на другом - обычно ложкообразный рудимент второго (нижнего) гнезда пыльника. Разросшийся в длинную нить связник (часть тычинки между гнездами пыльников) подвижно закреплен на очень короткой тычиночной нити. Полная редукция одного из гнезд пыльников у двух верхних тычинок имеет место еще у шлемника и железницы, но удлинения связника здесь не происходит.
Нектарники губоцветных являются производным основания плодолистиков. Наиболее обычный тип нектарника - это диск с 4 лопастями или зубцами. Каждая лопасть может выделять нектар, но эта способность находится в зависимости от степени развития самих лопастей и их проводящей системы. Насекомые находят нектар под завязью в нижней части венчика, однако при обильном выделении нектара им равномерно заполняется вся нижняя часть трубочки венчика и насекомому достаточно опустить хоботок в трубку, чтобы набрать много нектара. У шлемника сплошной нектароносный диск обычно замещен подковообразным нектарником с 3-5 неравными долями.
В строении гинецея всех губоцветных много общего. Он всегда образован двумя плодолистиками с числом гнезд, соответствующим числу плодолистиков. Однако каждое из гнезд делится ложной перегородкой пополам, вследствие чего завязь становится четырехлопастной, с одним семязачатком в каждой лопасти. Столбик у большинства губоцветных отходит от основания лопастей завязи (гинобазический), но в подсемействах живучковых (Ajugoideae) и простантеровых (Prostantheroideae) он обычно не вполне гинобазический или даже отходит почти от верхушки завязи, как в семействе вербеновых. У шлемника завязь не сидячая, как у остальных губоцветных, а расположена на ножке, образованной сильно суженной нижней частью гинецея.
Хотя цветки губоцветных обычно обоеполые, но во многих родах (например, мята, тимьян - Thymus) наряду с ними встречаются и женские цветки с рудиментарными тычинками, обычно имеющие более мелкий и бледно окрашенный венчик. Значительно реже встречаются мужские цветки с рудиментом гинецея (например, у некоторых видов котовника). Клейстогамные цветки с не выступающим из чашечки и обычно не опадающим венчиком можно видеть у обычного во многих районах СССР однолетнего сорняка яснотки стеблеобъемлющей (Lamium amplexicaule). Эти цветки обычно образуются в неблагоприятных климатических условиях: ранней весной или поздней осенью.
Плод губоцветных состоит из 4 односемянных и большей частью орешкообразных долей, имеющих очень различную форму. При плодах венчик обычно опадает (но остается у клейстогамных цветков и у некоторых родов подсемейства живучковых), а чашечка всегда остается и нередко разрастается (особенно у видов рода молуцелла (Molucella, см. рис. 215) и гименократер (Hymenocrater). Эндосперм в зрелых семенах обычно отсутствует, реже сохраняется, что является примитивной особенностью. Наиболее развит эндосперм у видов австралийского подсемейства простантеровых и у рода тетрахондра (Tetrachondra). Наружная оболочка долей плода нередко несет бугорки, сосочки или волоски,что связано со способом их распространения.
По новейшим данным, семейство губоцветных насчитывает около 200 родов и 3500 видов, распространенных почти по всему земному шару. Особенно многочисленны губоцветные в странах древнесредиземноморской флоры - от Канарских островов до Западных Гималаев, где они часто играют заметную роль в растительных группировках. Почти полностью отсутствуют губоцветные в Арктике и Антарктике. Очень мало губоцветных и в зоне тайги. Довольно богаты губоцветными горные районы тропиков, особенно Центральная и Южная Америка. В Австралии представлены главным образом эндемичные для этого континента роды подсемейства простантеровых (6 родов и около 100 видов). Еще беднее губоцветными Новая Зеландия, где имеются лишь по одному виду шлемника и мяты (оба эндемичных) и один из двух видов очень своеобразного рода тетрахондра (второй вид встречается в Патагонии). Род тетрахондра иногда выделяется в отдельное семейство. Относительно богаты губоцветными Гавайские острова с 2 эндемичными родами преимущественно тропического подсемейства празиевых.
Среди губоцветных преобладают нагорные и равнинные ксерофиты на сухих открытых местообитаниях, однако среди них немало и мезофильных лесных и луговых растений. В тропических дождевых лесах представлены лишь немногие виды. Настоящих водных растений среди губоцветных нет совсем, но имеется несколько родов, многие виды которых обитают по берегам водоемов и на болотах. Таковы, например, очень широко распространенные роды мята, зюзник, шлемник.
,
Взаимоотношения губоцветных с их опылителями-насекомыми (а в тропической и субтропической Америке также колибри) очень сложны и являются результатом длительной сопряженной эволюции. Виды родов с наиболее просто устроенными цветками, имеющими почти правильный венчик с короткой трубкой и 4 тычинки почти одинаковой длины (например, мята см. рис. 214) обычно опыляются мелкими перепончатокрылыми и мухами, так как нектар в них легко доступен. У большинства других губоцветных с хорошо выраженной двугубостью венчика тычинки и столбик прилегают к верхней губе, а нектар помещается в нижней части довольно длинной трубки. Опылителями таких цветков являются преимущественно перепончатокрылые и бабочки, реже крупные мухи из семейства журчалок. Посетители цветка прикасаются спинкой сначала к рыльцу, а затем к пыльникам и уносят на ней часть пыльцы. У яснотки и некоторых других родов высыпание пыльцы на спинку насекомого облегчается присутствием на пыльниках обращенных вниз волосков, за которые насекомое задевает. Замечательные приспособления типа рычага или шлагбаума, который открывает доступ к нектару лишь после попадания пыльцы на спинку насекомого, имеются у видов зопника и черноголовки (Prunella), однако наибольшего совершенства они достигают у видов шалфея, у которых пыльники двух имеющихся тычинок преобразованы в своеобразные подвижные рычажные устройства (см. выше). Опыляющее насекомое, чтобы достать нектар, просовывает голову в трубку венчика, отодвигает внутрь и кверху ложкообразно расширенный рудимент нижнего гнезда пыльника, а находящееся на другом конце удлиненного связника фертильное гнездо пыльника ударяет по спинке насекомого, высыпая на нее пыльцу (см. рис. 216). У американских губоцветиых из родов шалфей, шлемник, монарда и других нередки крупные красные цветки, опыляемые крупными ночными бабочками и колибри. Последние, как и бабочки из семейства бражников, парят возле цветков, высасывая клювом нектар и касаясь головкой расположенных под верхней губой или выступающих из венчика рылец и тычинок.
У некоторых губоцветных (особенно у родов подсемейства базиликовых) тычинки и столбик помещаются на нижней губе, так что насекомое, посещающее цветок (обычно бабочки), уносит пыльцу на нижней стороне брюшка. В других родах губоцветных тот же самый эффект (нижнее положение тычинок и столбика) достигается за счет перекручивания трубки венчика (верхняя губа становится как бы нижней губой), перекручивания цветоножки и сильно поникающего соцветия (например, у шалфея поникшего - S. nutans цветущие соцветия обращены вниз верхушкой). Возможность самоопыления цветков губоцветных нередко устраняется за счет более раннего созревания тычинок по сравнению с рыльцем (протандрии), однако во многих других случаях самоопыление вполне возможно. Следует отметить, что у многих губоцветных в привлечении насекомых и птиц принимают участие не только венчик, но и другие части цветка и соцветия. Так, у шалфея блестящего имеются ярко-красные чашечки, а у шалфея дубравного (S. nemorosa) - сине-фиолетовые прицветники.
Очень многие губоцветные распространяются с помощью ветра (анемохория). Единицами распространения - диаспорами - в этом случае обычно являются односемянные части дробного плода, парусность которых может увеличиваться, за счет их волосистости или образования крыловидных или хохолковидных выростов. Так, у распространенного в тропической Африке рода тиннея (Tinnea) плоды имеют хохолкообразные щитки. У анемохорных губоцветных стебли часто долго сохраняются в сухом состоянии, постепенно рассеивая плоды (даже в зимнее время). В других случаях, напротив, растопыренно разветвленные стебли с плодоносящими соцветиями легко обламываются у своего основания и перекатываются ветром по степи, постепенно разбрасывая плоды. Такими перекати-поле являются некоторые виды шалфея, зопника, котовника и др. Чем дольше плоды не выпадут из чашечек, тем на большее расстояние они перенесутся. Поэтому у многих губоцветных имеются приспособления для удерживания плодов в чашечке: кольцо волосков в ее зеве или загнутые внутрь зубцы.
У многих анемохорных губоцветных доли плода опадают вместе с чашечкой. Парусность в этих случаях достигается или за счет относительно длинных и нередко реснитчатых зубцов чашечки (например, у тимьяна - Thymus), или за счет сильного разрастания трубки чашечки и ее зубцов. Так, у молуцеллы (Molucella) трубка чашечки при плодах сильно увеличенная, перепончатая, ширококолокольчатая (см. рис. 214), а у зайцегуба, напротив, сильно разрастаются в ширину зубцы чашечки.
У некоторых видов отостегии (Otostegia) роль летучки выполняет сильно расширенная перепончатая верхняя губа чашечки, а у алжирского саккокаликса (Saccocalyx satureioides) чашечка при плодах пузыревидно вздутая с закрытым зубцами зевом, что позволяет находящемуся в ней плоду переноситься ветром иа большие расстояния.
Среди губоцветных немало видов, распространяющихся с помощью животных, причем приспособления к зоохории не менее разнообразны. Многие губоцветные имеют ослизняющиеся при смачивании оболочки долей плода и могут распространяться как эндозоохорно (с помощью поедающих плоды животных, преимущественно птиц), так и эпизоохорно (на шерсти и перьях, а также на ногах животных и человека). Большая эффективность эндозоохории достигается путем образования костянковидных долой плода с сочной мясистой оболочкой (у средиземноморского рода празиум). У рода хослундия (Hoslundia) из тропической Америки мясистой (ягодообразной) при плодах становится чашечка, зев которой закрыт зубцами. Эпизоохорно распространяются виды с клейкими или волосистыми долями плода. Во многих случаях для эпизоохорного распространения служат также опадающие вместе с плодами чашечки, жесткие волоски и жесткие оттопыренные в стороны зубцы, которые являются отличным приспособлением для заякоривания в шерсти животных.
Плоды некоторых губоцветных (особенно видов живучки и яснотки) имеют различным образом устроенные придатки, служащие пищей муравьям. Им свойствен так называемый мирмекохорный способ распространения. Бразильский вид хиптис Глазью (Н. glasiovii) вообще принадлежит к растениям «муравьелюбам» (мирмекофилам): во вздутых междоузлиях его стеблей постоянно находятся поселения особых муравьев.
Виды губоцветных, обитающих у берегов водоемов и на болотах (например, виды мяты и зюзника), имеют плавающие доли плода, приспособленные к распространению водными потоками, но отчасти и водными животными.
Система губоцветных еще далека от совершенства и находится в стадии разработки. Прежде всего еще не вполне ясна граница, отделяющая губоцветных от близкого, но более примитивного семейства вербеновых. Так, некоторые авторы предлагают относить к вербеновым 2 подсемейства губоцветных, сходные по строению гинецея с многими родами вербеновых,- простантеровые и живучковые; другие, напротив, предлагают перенести в губоцветные значительную часть семейства вербеновых. Согласно одной из последних систем семейства губоцветных, разработанной немецким ботаником X. Мельхиором (1964), оно делится иа 9 подсемейств. Первое место среди них занимает австралийское подсемейство простантеровые (Prostantheroideae), отличающееся относительно примитивным строением гинецея и семенами с эндоспермом, но имеющее довольно высокоспециализированное строение околоцветника. Далее следует подсемейство живучковых (Ajugoideae), у которого гинецей, как у простантеровых, но семена без эндосперма. Сюда относятся роды живучка, дубравник (Teucrium), аметистея (Amethystea) и др. В особое монотипное подсемейство розмариновые (Rosmarinoideae) выделяется род розмарин (Rosmarinus, табл. 55) с резко выраженным двугубым венчиком, 2 тычинками и семенами без эндосперма (см. рис. 214).
,
Следующее подсемейство - базиликовые (Ocimoideae), как и все последующие подсемейства, отличается от предыдущих подсемейств более специализированным гинецеем с ясно выраженным гинобазическим столбиком. Тычинок 4, редко 2. Представители этого подсемейства распространены почти исключительно в тропических и субтропических странах. Самый большой род хиптис (Hyptis) заключает более 350 видов, распространенных главным образом в Южной и Центральной Америке. К этому роду относятся самые высокие среди губоцветных деревья, произрастающие в лесах Бразилии. В род хиптис входят два хозяйственно важных вида: хиптис колосоносный (Н. spicigera), культивируемый для получения из его семян масла, похожего на кунжутное, и хиптис пахучий (H. suaveolens), или «сангура», дающий очень ароматичный лечебный чай. Род базилик (Ocimum) насчитывает до 150 видов, распространенных в тропических и субтропических странах, особенно в Африке. К этому роду относятся базилик благородный (О. basilicum), родом из тропической Азии, возделываемый во многих странах, в том числе в южных частях СССР, как пряное растение. В Китае культура этого растения известна с 500 г. до н. э. Другой известный представитель этого рода - базилик эвгенольный (O. gratissimum) - кустарник, родом из тропической Азии, культивируется, в частности, у нас в Грузии и в южных районах Краснодарского края как эфиромасличyое растение. Пользуется известностью также базилик священный (О. sanctum) - палеотропbческий кустарник, возделываемый в Индии b в других странах как культурное растение. Род шпороцветник (Plectranthus) включает около 250 видов, распространенных в тропических и субтропических странах Старого Света. Ряд видов этого рода достигает на севере Японии и южных районов Дальнего Востока. Наконец, следует упомяпуть палеотропический род колеус (табл. 55, около 150 видов). Некоторые виды, в том числе колеус съедобный (С. edulis), имеют крахмалистые клубневидно утолщенные корни и культивируются как пищевые растения в тропиках Старого Света. Многие виды декоративны, и некоторые из них выращивают в комнатах и садах. Индо-малайский вид колеус амбоинский (С. amboinicus) употребляют в качестве приправы к еде, а корyи индийского колеуса веттивериовидного (С. vettiverioides) идут на различные украшения. В подсемейство катофериевые (Catopherioideae) входит лишь один род катоферия (Catopheria, 3 вида), распространенный от Мексики до Колумбии. Виды катоферии - очень оригинальные по облику растения, характеризующиеся зародышем с сочным корешком, прилегающим к семядолям.
Подсемейство лавандовые (Lavanduloideae) также заключает только один род лаванда (Lavandula). Род лаванда, насчитывающий около 28 видов, распространен главным образом в Средиземноморье и Макаронезии, yо его ареал простирается до Сомали в Африке и до Индии. Сюда входят полукустарники и кустарники. Некоторые виды с древних времен используют для получения ценных эфирных масел. Лаванда узколистная (L. angustifolia) - кустарник до 1 м, а иногда до 2 м высотой, широко культивируется для получения ценных эфирных масел и очень популярен также как декоративное растение. Эфирные масла получают также из лаванды широколистной (L. latifolia) и некоторых других видов. Сухие цветки и листья лаванды долго сохраняют пряный запах, их используют для отпугивания моли. Следующее подсемейство - празиевые (Ргаsioideae) состоит из 6 родов, распространенных преимущественно в тропической Азии. Лишь один монотипный род празиум (Prasium) встречается в Средиземноморье от Португалии до Югославии. Для празиума, как и для других представителей подсемейства, характерны костянковидные доли плода.
К обширному подсемейству яснотковые (Lamioideae, у Мельхиора оно называется Stachyoideae) принадлежит подавляющее большинство внетропических губоцветных. Из представителей этого подсемейства следует прежде всего назвать род погостемон (Pogostemon) с около 40 видами, распространенными в Китае и в тропической Азии. К этому роду относится пачули (P. cablin) - происходящее из Филиппин очень ароматическое растение. Его широко культивируют в тропических странах для получения эфирного масла. Пачулевое масло обладает бактерицидными свойствами и широко используется в парфюмерии и медицине. К числу полезных представителей подсемейства относятся также 5 видов рода перилла (Реrillа), распространенных в Южной, Восточной и Юго-Восточной Азии. Перилла кустарниковая (Р. frutescens) культивируется в Восточной Азии в качество масличного и лекарственного растения, а перилла курчавая (P. frutеscens var. crispa) с темно-пурпуровыми, курчавыми листьями очень декоративна и в Китае и Японии выращивается как масличная, эфиро-масличиая и салатная культура. Еще большее значение имеет род мята (Mentha, около 25 видов в умеренной зоне северного полушария, в Южной Африке и в Австралии). Цветки у видов мяты почти актиноморфные, четырехчленные, с 4 почти одинаковыми тычинками. Некоторые виды мяты, особенно гибридная мята перечная (М. piperita), широко культивируют как ценные лекарственные и пищевые (в качестве приправы) растения. Мятные масла, содержащие наряду со многими другими компонентами значительное количество ментола, добавляют во многие лекарственные препараты, в конфеты и в зубную пасту. Иссоп лекарственный (Hyssopus officinalis) также культивируют как эфиромасличное, лекарственное и декоративное растение. Определенное значение имеют также виды рода душица (Origanum). Известно около 15-20 видов этого рода, распространенных в Европе, Средиземноморье и в умеренных областях Азии. Душицу обыкновенную (О. vulgare) используют как лекарственное растение, а листья употребляют как пряность и приправу в пищу и в ликеро-водочном производстве. Широко культивируют майоран (О. majorana), вместе с несколькими близкими видами иногда выделяемый в отдельный род Majorana. Родина майорана - Юго-Западная Азия и Северная Африка. Листья майорана используют в пищу как пряность к различным блюдам и для придания аромата уксусу и чаю. Из листьев и цветков добывают эфирное масло. Одним из наиболее известных представителей семейства является род тимьян (Thymus), насчитывающий от 35 до 400 видов в зависимости от точки зрения систематика на объем вида. Листья тимьяна содержат эфирные масла, главным образом тимол, используемый в медицине. Листья используют как пряность и в консервной и ликеро-водочной промышленности. Средиземноморский тимьян обыкновенный (Т. vulgaris) широко культивируется в умеренных и тропических странах. Используются также виды рода мелисса (Melissa, 5 видов в Евразии). Мелиссу лекарственную, или лимонную мяту (М. officinalis), культивируют как эфиромасличное, медоносное и пряное растение. Близок к мелиссе род чабер (Satureja), насчитывающий до 200 видов, распространенных в умеренных и субтропических областях. Чабер садовый (S. hortensis) культивируют как эфиромасличное растение. Его используют как пряность, в медицине и парфюмерии, а также для ароматизации ликеров и коньяков. Наконец, чабер горный (S. montana) разводят как декоративное растение.
Чистец (Stachys) - один из крупных родов подсемейства, насчитывающий около 300 видов, распространенных в умеренных, субтропических и тропических областях, отсутствующих, однако, в Австралии и Новой Зеландии. Некоторые виды чистеца играют заметную роль в сложении растительного покрова (табл. 55). Из числа полезных видов следует упомянуть так называемый китайский артишок (S. affinis), введенный в культуру в Китае и в настоящее время разводимый также в Японии, Бельгии и Франции в качестве овощного растения ради съедобных клубневидных корневищ. Несколько видов чистеца культивируются в качестве декоративных растений.
Шалфей - самый крупный род в семействе губоцветиых. Число видов шалфея достигает 700, и они широко распространены в умеренных, субтропических и тропических областях. Некоторые виды шалфея играют заметную роль в растительном покрове. Широко культивируется и всем хорошо известен шалфей лекарственный (S. officinalis). Очень популярным декоративным растением садов и парков стал бразильский шалфей блестящий с ярко-красной чашечкой и венчиком. Мексиканский вид шалфей наркотический (S. divinorum) содержит вещества, обладающие наркотическим действием, что было известно еще древним мексиканцам. В Перу священным цветком считался шалфей супротивноцветковый (S. oppositiflorа) - кустарник с красными цветками длиной 2,5-3 см.
Из других лекарственных представителей этого подсемейства укажем также пустырник (Leonurus cardiaca) - известное сердечное средство, кадило (Melittis melissophyllum), виды зизифоры (Ziziphora), зайцегуба.
Последнее место в системе Мельхиора занимает подсемейство шлемниковые (Scutellarioideae), наиболее специализированное по строению цветков. В это подсемейство входят только два рода - большой род шлемник (Scutellaria), насчитывающий около 300 видов, распространенный очень широко по земному шару (за исключением Южной Африки), и монотинный род салазария (Salazaria), распространенный в США и Мексике.
Более естественной по сравнению с системой Мельхиора является система губоцветных, предложенная в 1967 г. Р. Вундерлих. Она основывается главным образом на строении плодов и пыльцевых зерен, а в последнее время находит подтверждение и данными хемотаксономии. Вундерлих принимает только 6 подсемейств: простаптеровые, живучковые, шлемниковые, чистецовые, чаберовые (Saturejoideae) и катофериевые. Подсемейство празиевых Мельхиора она объединяет с чистецовыми, а подсемейства розмариновые и лавандовые - с выделяемым ею из яснотковых (чистецовых) подсемейством чаберовых. Базиликовые Мельхиора также присоединяются Вундерлих к чаберовым, но занимают в этом подсемействе обособленное положение. Несмотря на то что филогенетическая система Вундерлих имеет ряд достоинств, она также, несомненно, подвергнется дальнейшим изменениям.
Жизнь растений: в 6-ти томах. - М.: Просвещение. Под редакцией А. Л. Тахтаджяна, главный редактор чл.-кор. АН СССР, проф. А.А. Федоров . 1974 .
ЯСНОТКОВЫЕ ЯСНОТКОВЫЕ (Lamiaceae) , губоцветные (Labiatae), семейство двудольных цветковых р-ний. Одно- и многолетние травянистые растения, полукустарники, кустарники, изредка лианы и небольшие деревья. Листья супротивные или мутовчатые, простые, без прилистников. Цветки обоеполые, неправильные, часто двугу
В нашей статье мы хотим поговорить о семействе губоцветных. По последним данным, их насчитывается около двухсот родов - 3500 видов. Они распространены практически по всему миру. Не произрастают только в Антарктике и Артике. Мало их и в таежной зоне, они предпочитают тропические горные районы. Семейство Губоцветные - весьма своеобразный представитель флоры. Растения, которые относятся к нему, интересны прежде всего как поставщики лекарственного сырья натурального происхождения.
Внешний вид растений
Растения, которые входят в семейство Губоцветные, внешне имеют характерное строение. Их очень просто узнать по цветку, у которого присутствует длинная трубка и двугубый зев, напоминающий открытую пасть сказочного существа. Такой отличительный признак имеет только семейство Губоцветные. Соцветие подобного вида практически не встречается у других растений.
Что касается плодов, то они также имеют необычную форму. Плод семейства губоцветных состоит из четырех односемянных, в виде орехов, долек.
Также отличительной чертой является супротивное расположение цельных листьев. Стебли, как правило, имеют четырехгранную форму. Характеристика семейства Губоцветные будет неполной, если не упомянуть о сильном аромате, который свойственен большему числу растений. Феноменальный аромат объясняется присутствием на разных частях растений желез, которые выделяют сложные эфирные масла. Именно из-за присутствия этих масел растения семейства Губоцветные широко используются в качестве лекарственных, ароматических и О самых распространенных из них поговорим далее.
Семейство Губоцветные: представители
Представители данного семейства очень разнообразны. Большая их часть - это полукустарники и травы. Однако в субтропиках и тропиках очень распространены именно кустарники. Ярким представителем служит распространенный в Средиземноморье розмарин лекарственный. Это вечнозеленый кустарник с маленькими линейными листочками и фиолетовыми цветками.
Семейство губоцветных (фото приведено в статье) также представлено деревьями, но они встречаются только в тропиках. Некоторые из них достигают в высоту пятнадцати метров. Но, как правило, древесные губоцветные не вырастают более пяти метров.
Травянистые яснотковые
Трава семейства губоцветных - это прямостоячее растение, ему не требуется опора. Однако встречаются и стелющиеся виды (например, будра плющевидная). А у такого растения, как есть не только прямостоящие побеги, но и в пазухах листьев еще вырастают дугообразные отростки, укореняющиеся в земле своими верхушками (как усики у клубники).
Корневая система
Растения сохраняют свой основной корень в течение всей своей жизни. Иногда бывают случаи, когда он отмирает и его замещают придаточные корешки, которые отходят от самого стебля или от его ползучих побегов. Корневища характерны для большего числа губоцветных.
Гораздо реже встречаются растения с корневыми отпрысками, как у У прибрежных разновидностей, произрастающих на переувлажненных почвах, иногда корни имеют вид клубней, которые местные жители употребляют в пищу.
Строение листьев
Представители семейства имеют, как правило, цельные, иногда цельнокрайние листья. Но иногда встречаются голые виды, как, например, декоративный шалфей. В таком случае растение покрыто густым слоем волосков (чистец критский, железница крымская).
Цветки губоцветных
Как мы уже отмечали ранее, особое строение имеют и цветы. Семейство Губоцветные характеризуется тем, что последние у них, как правило, обоеполые. Они располагаются в пазухах листьев. Лишь у немногих видов встречаются одиночные цветки. Наиболее часто они собраны в соцветия по две штуки, которые образуют так называемые колоски. У некоторых разновидностей даже имеются колючки, которые призваны защищать растение от поедания животными. Такие виды встречаются в горных районах Средней Азии (зайцегуб).
Как правило, и чашечка, и венчик губоцветных состоят из пяти листочков, сросшихся в трубку. Вообще, чашечка бывает самых разных форм: колокольчатой, трубчатой, ворончатой, шаровидной. Ее видоизменения связаны с попыткой приспособиться к распространению плодов. Чашечка может менять цвет, становясь очень яркой, для привлечения внимания птиц и животных, может разрастаться, увеличивая парусность, для распространения семян ветром.
Семейство Губоцветные - растения, которые имеют в каждом цветочке по четыре тычинки, закрепленных на трубочке венчика. У некоторых тропических видов тычинки срастаются вместе. Ниже их места расположения имеется волосистое кольцо, которое призвано защищать нектар.
Пыльники губоцветных могут быть самых разных форм. Все зависит от степени приспособленности растения к опылению. Иногда встречаются настоящие сложные «механизмы», как у железницы и шлемника.
Хоть для губоцветных более характерны обоеполые цветки, у многих представителей наряду с ними есть и женские с тычинками. Намного реже можно встретить мужские цветки. Примером такого растения является травянистое растение семейства губоцветных яснотка стеблеобъемлющая (сорняк).
Плоды
Семейство Губоцветные, представители которого рассматриваются нами, отличается тем, что все растения имеют характерное строение плода. Как правило, он состоит из четырех долей, в каждой из которых находится по одному семени. Сама форма плода может быть очень разной. Венчик при плодах обычно опадает, а вот чашечка непременно остается и разрастается. В зрелых семенах эндосперм отсутствует.
Внешняя оболочка долей часто имеет бугристую форму, что способствует распространению семян.
Места произрастания
Представители такого вида флоры, как семейство Губоцветные (Яснотковые), распространены по всему миру. Особенно их много в странах со средиземноморской флорой (начиная от Канарских островов и до Западных Гималаев). А вот в тайге такие растения практически отсутствуют. Рай для губоцветных - тропические горные районы. Особенно их много в Южной и Центральной Америке. В Австралии встречаются только эндемики этого континента, всего около ста видов. Еще беднее оказалась Новая Зеландия, там присутствует только один вид мяты и одна разновидность шлемника, а также один представитель рода Тетрахондра. Зато на Гавайских островах довольно много губоцветных, они представлены двумя эндемическими родами.
Растения данного семейства предпочитают расти на открытых горных или равнинных местах. Для них подходят более сухие грунты. Среди них очень мало луговых и лесных растений. В дождевых тропических лесах способны выживать только немногие представители, поскольку губоцветные не выносят излишней влаги. Что касается настоящих водных разновидностей, то их вообще не существует в природе. Есть только несколько родов, некоторые виды которых растут на берегах болот и водоемов. Примером может служить многолетнее травянистое растение семейства губоцветных - мята, а также шлемник и зюзник.
Опыление растений насекомыми
Отношения губоцветных с насекомыми, которые их опыляют, достаточно сложные и являются результатом длительного процесса эволюции. Те растения данного семейства, что имеют наиболее просто устроенные цветочки, опыляются мухами и перепончатокрылыми, поскольку нектар из них достать не представляет особого труда.
У губоцветных, имеющих более сложное нектар не так-то и просто добыть. Он находится на дне длинной трубки. Опылением таких видов занимаются бабочки и перепончатокрылые, очень редко большие мухи-журчалки.
Уникальным приспособлением в виде рычага, который дает возможность добраться до нектара насекомому, обладает шалфей. Насекомым нужно изловчиться, чтобы достать то, зачем они прилетели. В субтропических и тропических регионах Америки растения опыляют маленькие птички колибри. Таким же способом, как и колибри, действуют бабочки семейства бражников. Они парят вокруг цветочков и при этом высасывают нектар клювом, касаясь головой тычинок.
Некоторые растения из семейства губоцветных имеют такое строение цветка, что насекомое, садясь на него, цепляет пыльцу своим брюшком и затем уносит ее. Очень редкие разновидности могут самоопыляться.
Хочется отметить, что у губоцветных есть свои способы привлечения внимания насекомых, например, в виде отдельных ярких частей цветка.
Приспособленность растений к распространению
Что касается размножения, то преобладающее большинство распространяется при помощи ветра. В самом процессе участвуют односемянные доли плода, парусность которого увеличивается за счет волосков или крыловидных выростов. В Африке очень распространен род Тиннея. Так вот, его плоды вооружены хохолкообразными щитками, которые помогают распространению семян.
Некоторые разновидности долгое время сохраняют стебли в сухом виде, постепенно семена сами рассеваются под действием ветров. У иных же, наоборот, они очень быстро обламываются у земли и ветрами переносятся по степям, рассевая при этом плоды. Такой эффект перекати-поле характерен для некоторых разновидностей шалфея, котовника, зопника. Чем дольше плоды будут находиться в чашечке, тем на более дальнее расстояние они перенесутся. Именно по этой причине у многих губоцветных имеются надежные приспособления для удержания семян.
Иные виды, наоборот, устроены таким образом, что плод падает вместе с чашечкой и за счет нее имеет большую парусность, которая позволяет ему распространяться на дальние расстояния.
Среди губоцветных есть немало растений, которые распространяются при помощи живых существ. При этом они имеют не менее интересные приспособления, которые им помогают в этом. Некоторые из них поедаются птицами и животными, другие цепляются отростками за их шерсть и одежду человека. Каждый вид нашел свой путь распространения.
В тропиках некоторые плоды покрыты мясистой оболочкой (в виде ягодки), что привлекает зверей и птиц, другие же смазаны клейким веществом, позволяющим прилипнуть им к шерсти или оперению.
А вот приспособленность к распространению некоторых видов вообще поражает. Например, некоторые разновидности яснотки и живучки имеют вещества, которые служат пищей для муравьев, именно с их помощью и распространяются плоды. Бразильское растение хиптис устроено таким образом, что в его междоузлиях вообще всегда обитают колонии муравьев.
Те губоцветные, что выбрали местом своего обитания побережье рек и болот, имеют плавающие части плода, и потому распространяются водным способом, иногда и при помощи животных.
Применение
Трава, кустарник, лиана, полукустарник семейства губоцветных - все это разнообразные формы одного и того же семейства. Многие из них являются эфирномасличными культурами, а потому представляют особый интерес для человека. К таким растениям относятся: базилик, шалфей, шандра, дубровник, розмарин, лаванда.
Лаванда - это полукустарник семейства губоцветных. Известно более двадцати пяти ее видов. Во многих странах ее возделывают исключительно для получения уникального эфирного масла. А некоторые виды этого интересного растения представляют интерес как декоративный лекарственный медонос.
Базилик также относится к кустарниковым и полукустарниковым растениям. В субтропиках и тропиках произрастает около 150 его видов, некоторые из них дают ценное эфирное масло. Кроме того, базилик широко используют в качестве приправы во многих кухнях мира.
Знаменитый шалфей также имеет множество разновидностей, некоторые из них являются медоносами и источником получения ароматического масла. В России он распространен в степной зоне.
Перилла - это представитель однолетних губоцветных. Это исключительно масличная культура. Ее выращивают в Азии и Японии, Китае и Северной Корее, и даже на Дальнем Востоке. Из семян получают пищевое и техническое масло. Кроме того, некоторые виды имеют декоративное применение, а также являются прекрасными медоносами.
Лекарственные растения семейства губоцветных
Издавна яснотковые в народе ценились за свои лечебные свойства. И в настоящее время активно используют их целительные вещества. В наших регионах наиболее известны такие разновидности: мята, пустырник, шалфей, зюзник, и т. д.
Шалфей - это не только но еще и лекарственное растение семейства губоцветных. Его активно используют для орошения и полосканий горла и ротовой полости.
Базилик хорош при потере аппетита, при запорах и метеоризме. Иногда его применяют как полоскание для горла, для компрессов при загноившихся ранах.
Душица - также лекарственное растение семейства губоцветных, используется для лечения кишечных и желудочных заболеваний, а также при бронхитах. Следует заметить, что именно масло душицы является частью многих мазей для растираний, которые хороши при ревматизме. Также растение употребляют в качестве пряности, например, в итальянской кухне для приготовления знаменитой пиццы. Эфирное масло этого растения замечательно. А в индийской медицине душица применяется не только для лечения желудочных болезней, но и при нервных расстройствах.
Знакомая нам с детства перечная мята является частью многих чайных сборов, которые помогают при лечении печени, желчного пузыря, кишечника и желудка. Вообще ее можно встретить во многих сборах. Кроме того, в домашних условиях ее заваривают как простой чай, поскольку напиток получается очень ароматным, приятным и имеет седативное действие.
Дубровник обыкновенный применяют как желудочное лекарство при заболеваниях желчного пузыря и кишечника. Кроме того, он имеет свойство возбуждать аппетит и помогает при бронхитах.
Майоран нам знаком как приправа. Он улучшает аппетит, стимулируя образование желчи и желудочного сока. Его настойка имеет антисептическое, спазмолитическое, мочегонное и общетонизирующее действие. Майоран применяют при гастритах и хронических холециститах, метеоризме и головных болях, нарушениях цикла, бессоннице, рвоте.
Пикульник хорош при кашле и разных легочных заболеваниях. Кроме того, он обладает кровоочистительными свойствами, применяется при заболеваниях желчного пузыря и печени.
Почечный чай - замечательное мочегонное средство, которое широко применяют при заболеваниях мочевыводящих путей.
Чабер благотворно влияет на желудок, способствуя выделению сока, поэтому его применяют для возбуждения аппетита. Заваренный в виде чая, он используется при насморках и кашлях.
Тимьян используют в медицине как отхаркивающее средство.
В рамках статьи мы рассказали лишь о некоторых лекарственных растениях семейства губоцветных. На самом деле их очень много, обо всех поведать невозможно. Но о том, что они на самом деле помогают при разного рода заболеваниях, свидетельствует многолетняя практика их применения в разных странах мира.
Губоцветные вокруг нас
Хочется заметить, что растения семейства губоцветных растут не только в условиях дикой природы. Вы удивитесь, но среди культурных растений на наших клумбах достаточно представителей этой группы, например те же сальвии, радующие нас своими яркими цветками до прихода морозов.
Кроме того, некоторые цветы, растущие на подоконниках наших квартир, также относятся к губоцветным: колеус, шведский плющ, мольное дерево. Хозяйкам они давно полюбились за свою неприхотливость и яркую окраску. Они не нуждаются в особом уходе, зато всегда радуют своей красотой. В летний период растения отлично растут на балконе, а зимой в квартире. Они предпочитают яркое освещение, поэтому ставить их нужно на южную строну.
Что касается полива, то он не должен быть очень частым. А в зимний период, скорее, редким. Как мы говорили ранее, губоцветные не переносят лишней влаги, это касается и домашних представителей.
В последнее время очень модным стало такое растение, как розмарин. Его не только используют в качестве приправы, но и выращивают в домашних условиях. Правда, растение не любит высоких температур и сухого воздуха центрального отопления (может даже сбросить листья и начать усыхать). Оно имеет неплохой декоративный вид, особенно интересным становится во время цветения, когда весь покрывается мелкими синими или голубыми цветочками. На летний период некоторые хозяйки даже высаживают такие домашние растения в открытый грунт, там они себя прекрасно чувствуют. Но перед приходом холодов розмарин необходимо вернуть вновь домой, поскольку он не перенесет морозов. Данная культура хороша для домашнего применения не только из-за декоративного вида. Растение издает приятный аромат, очищая воздух, поскольку обладает бактерицидными свойствами.
Вместо послесловия
Среди растений, окружающих нас в повседневной жизни, немало тех, что принадлежат к семейству губоцветных. Как мы убедились, они не только украшают наши дома и цветники, но и широко применяются в медицине и парфюмерии. А ведь о существовании многих из них мы даже не задумывались, тем более не знали, что они принадлежат к столь обширному семейству.
Семейство ГУБОЦВЕТНЫЕ (LAMIACEAE)
Представители этого семейства легко узнаются уже по строению венчика цветков, имеющего длинную трубку и двугубый зев, напоминающий разинутую пасть сказочного животного. Хотя сходный по облику венчик имеется еще в нескольких семействах цветковых растений (например, у многих норичниковых), для губоцветных он особенно характерен. Не менее своеобразен и плод губоцветных, состоящий из 4 односемянных орешкообразных или редко костянковидных долей, тогда как у сходных с ними по строению венчика норичниковых плод — обычно многосемянная коробочка. К числу важных отличительных особенностей губоцветных принадлежат также супротивные (очень редко мутовчатые), обычно цельные листья без прилистников и обычно четырехгранные стебли. Очень существен ароматический запах, свойственный большинству видов губоцветных, который определяется присутствием на всех или на некоторых частях растения желёзок, выделяющих эфирные масла сложного состава (в них входят ароматические спирты, фенолы, терпены, альдегиды и другие органические соединения). Именно присутствием этих масел в значительной степени определяется практическое использование губоцветных в качестве технических, лекарственных и ароматических растений.
Большинство губоцветных — травы и полукустарники. Однако среди них, особенно в тропиках и субтропиках, много и кустарников, примером которых может служить распространенный в Средиземноморской флористической области розмарин лекарственный (Rosmarinus officinalis) — вечнозелёный кустарник с мелкими линейными листьями и сине-фиолетовыми (до почти белых) цветками. Губоцветные — деревья имеются только в тропиках, но, в отличие от близкого преимущественно древесного семейства вербеновых, ими являются лишь немногие виды двух американских родов: хиптис (Hyptis) и левкосцептрум (Leucosceptrum). «Чемпион» по высоте среди них — бразильский вид хиптис перепончатый (Н. membranacea), достигающий в высоту 12—15 м, в то время как другие древесные губоцветные обычно не достигают высоты 5 м. В тропиках встречаются и немногие лианы, к которым принадлежат лишь американский род салазария (Salazaria), некоторые виды шлемника (Scutellaria) и гавайского рода стеногина (Stenogyne).
Стебли травянистых губоцветных обычно прямостоячие и не нуждаются в опоре, хотя имеются виды со стелющимися по земле и укореняющимися в узлах стеблями (например, будра плющелистная — Glechoma hederacea). У живучки ползучей (Ajuga reptans), кроме прямостоячих репродуктивных побегов, из пазух листьев розетки образуются дугообразные направленные к земле и укореняющиеся верхушками вегетативные побеги, подобные усам земляники. Хорошо развитая розетка прикорневых листьев, сохраняющаяся во время циетения растения, имеется у многих травянистых губоцветных (например, у некоторых шалфеев — Salvia). Главный корень часто сохраняется в течение всей жизни растения, реже отмирает и замещается придаточными корнями, отходящими или от основания стебля или от отходящих от него ползучих подземных побегов — корневищ, свойственных многим видам губоцветных. Довольно редко среди губоцветных встречаются виды с корневыми отпрысками, например живучка женевская (Ajuga gennevensis). У многих прибрежных видов, обитающих на переувлажненных местообитаниях, в корневищах образуются воздухоносные полости или участки воздухоносной ткани. Некоторые губоцветные имеют клубневидно утолщенные корни, в тропических странах употребляемые в пищу. При супротивном расположении листьев близлежащие пары крестообразно чередуются друг с другом. Следствием этого и является четырехгранность стеблей губоцветных, причем грани могут быть плоскими, выпуклыми или вогнутыми. Видов, имеющих мутовчатое расположение листьев, среди губоцветных немного. К ним принадлежит, в частности, оригинальный австралийский кустарник вестриигия (Westringia) с мелкими цельнокрайними листьями, расположенными мутовками по 3—6. Очередное листорасположение отмечено только у первых листьев проростков в родах зопник (Phlomis) и буквица (Betonica).
Листья губоцветных обычно цельные и часто цельнокрайние, хотя встречаются и перисто-раздельные (например, у шалфея скабиозолистного — Salvia scabiosifolia). Известны как голые или почти голые виды, например декоративный шалфей блестящий (S. splendens), так и виды, густо покрытые волосками. Среди последних такие средиземноморские виды, как чистец критский (Stachys cretica) и железница крымская (Sideritis taurica), не уступают по красоте знаменитому альпийскому эдельвейсу. Из волосков наиболее распространены многоклеточные простые волоски. Головчатые волоски, головка которых функционирует как желёзка, вырабатывающая эфирное масло, встречаются у многих губоцветных. Обычно пятичленные и, как правило, обоеполые цветки губоцветных располагаются в пазухах неизмененных или видоизмененных в прицветники листьев. Лишь в немногих случаях (например, у видов шлемника) они одиночные, обычно же цветки образуют так называемые ложные мутовки, составленные двумя супротивными соцветиями, несущими прицветнички. Оси всех первичных соцветий, часто сильно укороченные и составляющие их цветки, кажутся расположенными непосредственно в листовых пазухах, образуя колосовидные соцветия. Иногда ось общего соцветия сильно укорочена и все соцветие становится головкообразным, например у зизифоры головчатой (Ziziphora capitata) и у большого американского рода хиптис. У нередкого в горных районах Средней Азии зайцегуба (Lagochilus) расположенные при основании ложных мутовок прицветнички видоизменены в мощные колючки, защищающие растение от поедания травоядными животными. У некоторых других губоцветных в колючки видоизменяются прицветники или верхние листья, а иногда зубцы листьев.
И чашечка, и венчик губоцветных обычно образованы 5 сросшимися своей основной частью в трубку листочками. Лишь немногие роды, например западносредиземноморский род преслия (Preslia), имеют четырехчленный околоцветник. Чашечка губоцветных может иметь самую разнообразную форму: трубчатую, колокольчатую, ворончатую, шаровидную, а в зеве может быть как двугубой без каких-либо зубцов, так и 5(4)-зубчатой с зубцами одинаковой или различной длины (в последнем случае чашечка также более или менее двугубая). В отличие от венчика, видоизменения которого связаны с приспособлением к опылению, видоизменения чашечки обычно связаны с приспособлением к распространению плодов. Нередко (например, у пустырника) зубцы чашечки имеют вид отогнутых в сторону колючек. Иногда вся чашечка или ее зубцы сильно разрастаются, увеличивая парусность чашечки при распространении семян с помощью ветра, или чашечка становится ярко окрашенной, играя роль в привлечении насекомых или птиц-опылителей, например, ярко-красная чашечка шалфея блестящего. Очень оригинальна чашечка у большого (около 300 видов), почти космополитного рода шлемник. Она здесь двугубая с цельно-крайними губами и после созревания плода распадается на 2 части, имеющие вид створок: нижнюю остающуюся и верхнюю опадающую. Верхняя часть чашечки у многих видов этого рода несет еще поперечную чешуевидную складку — щиток. Шлемник имеет еще ряд особенностей, отличающих этот род от других родов губоцветных (в том числе отсутствие эфиромас-личных желёзок), и не случайно некоторые авторы даже предлагали выделить его в особое семейство шлемниковые (Scutellariaceae).
Как уже упоминалось выше, венчик губоцветных обычно разделен на две губы, из которых верхняя образована 2, а нижняя — 3 лепестками. Верхняя губа может быть плоской или выпуклой, иногда она бывает цельнокрайней, так что никаких следов присутствия 2 лепестков не обнаруживается. Нижняя губа почти всегда более крупная (посадочная площадка для опылителей), трехлопастная с более крупной и нередко, в свою очередь, двулопастной средней лопастью.
Плод губоцветных состоит из 4 односемянных и большей частью орешкообразных долей, имеющих очень различную форму. При плодах венчик обычно опадает (но остается у клейстогамных цветков и у некоторых родов подсемейства живучковых), а чашечка всегда остается и нередко разрастается (особенно у видов рода молуцелла (Molucella) и гименократер (Hymenocrater). Эндосперм в зрелых семенах обычно отсутствует, реже сохраняется, что является примитивной особенностью. Наиболее развит эндосперм у видов австралийского подсемейства простантеровых и у рода тетрахондра (Tetrachondra). Наружная оболочка долей плода нередко несет бугорки, сосочки или волоски,что связано со способом их распространения.
По новейшим данным, семейство губоцветных насчитывает около 200 родов и 3500 видов, распространенных почти по всему земному шару. Особенно многочисленны губоцветные в странах древнесредиземноморской флоры -от Канарских островов до Западных Гималаев, где они часто играют заметную роль в растительных группировках. Почти полностью отсутствуют губоцветные в Арктике и Антарктике. Очень мало губоцветных и в зоне тайги. Довольно богаты губоцветными горные районы тропиков, особенно Центральная и Южная Америка. В Австралии представлены главным образом эндемичные для этого континента роды подсемейства простантеровых (6 родов и около 100 видов). Еще беднее губоцветными Новая Зеландия, где имеются лишь по одному виду шлемника и мяты (оба эндемичных) и один из двух видов очень своеобразного рода тетрахондра (второй вид встречается в Патагонии). Род тетрахондра иногда выделяется в отдельное семейство. Относительно богаты губоцветными Гавайские острова с 2 эндемичными родами преимущественно тропического подсемейства празиевых. Среди губоцветных преобладают нагорные и равнинные ксерофиты на сухих открытых местообитаниях, однако среди них немало и мезофильных лесных и луговых растений. В тропических дождевых лесах представлены лишь немногие виды. Настоящих водных растений среди губоцветных нет совсем, но имеется несколько родов, многие виды которых обитают по берегам водоемов и на болотах. Таковы, например, очень широко распространенные роды мята, зюзник, шлемник. Взаимоотношения губоцветных с их опылителями-насекомыми (а в тропической и субтропической Америке также колибри) очень сложны и являются результатом длительной сопряженной эволюции. Виды родов с наиболее просто устроенными цветками, имеющими почти правильный венчик с короткой трубкой и 4 тычинки почти одинаковой длины (например, мята) обычно опыляются мелкими перепончатокрылыми и мухами, так как нектар в них легко доступен. У большинства других губоцветных с хорошо выраженной двугубостыо венчика тычинки и столбик прилегают к верхней губе, а нектар помещается в нижней части довольно длинной трубки. Опылителями таких цветков являются преимущественно перепончатокрылые и бабочки, реже крупные мухи из семейства журчалок. Посетители цветка прикасаются спинкой сначала к рыльцу, а затем к пыльникам и уносят на ней часть пыльцы. У яснотки и некоторых других родов высылание пыльцы на спинку насекомого облегчается присутствием на пыльниках обращенных вниз волосков, за которые насекомое задевает. Замечательные приспособления типа рычага или шлагбаума, который открывает доступ к кектару лишь после попадания пыльцы на спинку насекомого, имеются у видов зопника и Черноголовки (Prunella), однако наибольшего совершенства они достигают у видов шалфея, у которых пыльники двух имеющихся тычинок преобразованы в своеобразные подвижные рычажные устройства. Опыляющее насекомое, чтобы достать нектар, просовывает голову в трубку венчика, отодвигает внутрь и кверху ложкообразно расширенный рудимент нижнего гнезда пыльника, а находящееся на другом конце удлиненного связника фертильное гнездо пыльника ударяет по спинке насекомого, высыпая на нее пыльцу. У американских губоцветных из родов шалфей, шлемник, монарда и других нередки крупные красные цветки, опыляемые крупными ночными бабочками и колибри. Последние, как и бабочки из семейства бражников, парят возле цветков, высасывая клювом нектар и касаясь головкой расположенных под верхней губой или выступающих из венчика рылец и тычинок.
У некоторых губоцветных (особенно у родов подсемейства базиликовых) тычинки и столбик помещаются на нижней губе, так что насекомое, посещающее цветок (обычно бабочки), уносит пыльцу на нижней стороне брюшка. В других родах губоцветных тот же самый эффект (нижнее положение тычинок и столбика) достигается за счет перекручивания трубки венчика (верхняя губа становится как бы нижней губой), перекручивания цветоножки и сильно поникающего соцветия (например, у шалфея поникшего — S. nutans цветущие соцветия обращены вниз верхушкой). Возможность самоопыления цветков губоцветных нередко устраняется за счет более раннего созревания тычинок по сравнению с рыльцем (протандрии), однако во многих других случаях самоопыление вполне возможно. Следует отметить, что у многих губоцветных в привлечении насекомых и птиц принимают участие не только венчик, но и другие части цветка и соцветия. Так, у шалфея блестящего имеются ярко-красные чашечки, а у шалфея дубравного (S. nemorosa) — сине-фиолетовые прицветники.
Очень многие губоцветные распространяются с помощью ветра (анемохория). Единицами распространения — диаспорами — в этом случае обычно являются односемянные части дробного плода, парусность которых может увеличиваться за счет их волосистости или образования крыловидных или хохолковидных выростов. Так, у распространенного в тропической Африке рода тиннея (Tinnea) плоды имеют хохолкообразные щитки. У анемохорных губоцветных стебли часто долго сохраняются в сухом состоянии, постепенно рассеивая плоды (даже в зимнее время). В других случаях, напротив, растопыренно разветвленные стебли с плодоносящими соцветиями легко обламываются у своего основания и перекатываются ветром по степи, постепенно разбрасывая плоды. Такими перекати-поле являются некоторые виды шалфея, зопника, котовника и др. Чем дольше плоды не выпадут из чашечек, тем на большее расстояние они перенесутся. Поэтому у многих губоцветных имеются приспособления для удерживания плодов в чашечке: кольцо волосков в ее зеве или загнутые внутрь зубцы. У многих анемохорных губоцветных доли плода опадают вместе с чашечкой. Парусность в этих случаях достигается или за счет относительно длинных и нередко реснитчатых зубцов чашечки (например, у тимьяна - Thymus), или за счет сильного разрастания трубки чашечки и ее зубцов. Так, у молуцеллы (Molucella) трубка чашечки при плодах сильно увеличенная, перепончатая, ширококолокольчатая, а у зайцегуба, напротив, сильно разрастаются в ширину зубцы чашечки. У некоторых видов отостегии (Otostegia) роль летучки выполняет сильно расширенная перепончатая верхняя губа чашечки, а у алжирского саккокаликса (Saccocalyx satureioides) чашечка при плодах пузыревидно вздутая с закрытым зубцами зевом, что позволяет находящемуся в ней плоду переноситься ветром на большие расстояния.
Среди губоцветных немало видов, распространяющихся с помощью животных, причем приспособления к зоохории не менее разнообразны. Многие губоцветные имеют ослизняющиеся при смачивании оболочки долей плода и могут распространяться как эндозоохорно (с помощью поедающих плоды животных, преимущественно птиц), так и эпизоохорно (на шерсти и перьях, а также на ногах животных и человека). Большая эффективность эндозоохории достигается путем образования костянковидных долей плода с сочной мясистой оболочкой (у средиземноморского рода празиум). У рода хослундия (Hoslundia) из тропической Америки мясистой (ягодообразной) при плодах становится чашечка, зев которой закрыт зубцами. Эпизоохорно распространяются виды с клейкими или волосистыми долями плода. Во многих случаях для эпизоохорного распространения служат также опадающие вместе с плодами чашечки, жесткие волоски и жесткие оттопыренные в стороны зубцы, которые являются отличным приспособлением для заякоривания в шерсти животных.
Плоды некоторых губоцветных (особенно видов живучки и яснотки) имеют различным образом устроенные придатки, служащие пищей муравьям. Им свойствен так называемый мирмекохорный способ распространения. Бразильский вид хиптис Глазъю (Н. glasiovii) вообще принадлежит к растениям «муравьелюбам» (мирмекофилам): во вздутых междоузлиях его стеблей постоянно находятся поселения особых муравьев. Виды губоцветных, обитающих у берегов водоемов и на болотах (например, виды мяты и зюзника), имеют плавающие доли плода, приспособленные к распространению водными потоками, но отчасти и водными животными. внетропических губоцветных. Из представителей этого подсемейства следует прежде всего назвать род погостемон (Pogostemon) с около 40 видами, распространенными в Китае и в тропической Азии. К числу полезных представителей подсемейства относятся также 5 видов рода перилла (Perilla), распространенных в Южной, Восточной и Юго-Восточной Азии. Еще большее значение имеет род мята (Mentha, около 25 видов в умеренной зоне северного полушария, в Южной Африке и в Австралии). Цветки у видов мяты почти актиноморфные, четырехчленные, с 4 почти одинаковыми тычинками. Иссоп лекарственный (Hyssopus officinalis) также культивируют как эфиромасличиое, лекарственное и декоративное растение. Определенное значение имеют также виды рода душица (Origanum). Известно около 15—20 видов этого рода, распространенных в Европе, Средиземноморье и в умеренных областях Азии. Одним из наиболее известных представителей семейства является род тимьян (Thymus), насчитывающий от 35 до 400 видов в зависимости от точки зрения систематика на объем вида. Листья тимьяна содержат эфирные масла, главным образом тимол, используемый в медицине. Листья используют как пряность и в консервной и ликеро-водочной промышленности. Средиземноморский тимьян обыкновенный (Т. vulgaris) широко культивируется в умеренных и тропических странах. Используются также виды рода мелисса (Melissa, 5 видов в Евразии). Мелиссу лекарственную, или лимонную мяту (М. officinalis), культивируют как эфиромасличное, медоносное и пряное растение. Близок к мелиссе род чабер (Satureja), насчитывающий до 200 видов, распространенных в умеренных и субтропических областях. Чабер садовый (S. hortensis) культивируют как эфиромасличное растение. Его используют как пряность, в медицине и парфюмерии, а также для ароматизации ликеров и коньяков. Наконец, чабер горный (S. montana) разводят как декоративное растение. Чистец (Stachys) — один из крупных родов подсемейства, насчитывающий около 300 видов, распространенных в умеренных, субтропических и тропических областях, отсутствующих, однако, в Австралии и Новой Зеландии. Некоторые виды чистеца играют заметную роль в сложении растительного покрова. Шалфей — самый крупный род в семействе губоцветных. Число видов шалфея достигает 700, и они широко распространены в умеренных, субтропических и тропических областях. Некоторые виды шалфея играют заметную роль в растительном покрове. Широко культивируется и всем хорошо известен шалфей лекарственный (S. officinalis). Очень популярным декоративным растением садов и парков стал бразильский шалфей блестящий с ярко-красной чашечкой и венчиком. Мексиканский вид шалфей наркотический (S. divinorum) содержит вещества, обладающие наркотическим действием, что было известно еще древним мексиканцам. В Перу священным цветком считался шалфей супротивноцветковый (S. oppositiflo-га) — кустарник с красными цветками длиной 2,5—3 см. Из других лекарственных представителей этого подсемейства укажем также пустырник (Leonurus cardiaca) — известное сердечное средство, кадило (Melittis melissophyllum), виды зизифоры (Ziziphora), зайцегуба. Последнее место в системе Мельхиора занимает подсемейство шлемниковые (Scutellarioi-deae), наиболее специализированное по строению Система губоцветных еще далека от совершенства и находится в стадии разработки. Прежде всего еще не вполне ясна граница, отделяющая губоцветных от близкого, но более примитивного семейства вербеновых. Так, некоторые авторы предлагают относить к вербеновым 2 подсемейства губоцветных, сходные по строению гинецея с многими родами вербеновых,— простантеровые и живучковые; другие, напротив, предлагают перенести в губоцветные значительную часть семейства вербеновых. Согласно одной из последних систем семейства губоцветных, разработанной немецким ботаником X. Мельхиором (1964), оно делится на 9 подсемейств. Первое место среди них занимает австралийское подсемейство простантеровые (Prostantheroideae), отличающееся относительно примитивным строением гинецея и семенами с эндоспермом, но имеющее довольно высокоспециализированное строение околоцветника. Далее следует подсемейство живучковых (Ajugoideae), у которого гинецей, как у простанте-ровых, но семена без эндосперма. Сюда относятся роды живучка, дубравник (Teucrium), аметистея (Amethystea) и др. В особое монотипное подсемейство розмариновые (Rosmarinoideae) выделяется род розмарин (Rosmarinus) с резко выраженным двугубым венчиком, 2 тычинками и семенами без эндосперма.
Следующее подсемейство - базиликовые (Ocimoideae) , как и все последующие подсемейства, отличается от предыдущих подсемейств более специализированным гинецеем с ясно выраженным гинобазическим столбиком. Тычинок 4, редко 2. Представители этого подсемейства распространены почти исключительно в тропических и субтропических странах. Самый большой род хиптис (Hyptis) заключает более 350 видов, распространенных главным образом в Южной и Центральной Америке. К этому роду относятся самые высокие среди губоцветных деревья, произрастающие в лесах Бразилии. В род хиптис входят два хозяйственно важных вида: хиптис колосоносный (Н. spicigera), культивируемый для получения из его семян масла, похожего на кунжутное, и хиптис пахучий (Н. suaveolens), или «сангура», дающий очень ароматичный лечебный чай. Род базилик (Ocimum) насчитывает до 150 видов, распространенных в тропических и субтропических странах, особенно в Африке. К этому роду относятся базилик благородный (О. basilicum), родом из тропической Азии, возделываемый во многих странах, в том числе в южных частях бывшего СССР, как пряное растение. В Китае культура этого растения известна с 500 г. до н. э. Другой известный представитель этого рода — базилик эвгенолъный (О. gratissimum) — кустарник, родом из тропической Азии, культивируется, в частности, у нас в Грузии и в южных районах Краснодарского края как эфиромасличное растение. Пользуется известностью также базилик священный (О. sanctum) — палеотропиче-ский кустарник, возделываемый в Индии и в других странах как культурное растение. Род шпороцветник (Plectranthus) включает около 250 видов, распространенных в тропических и субтропических странах Старого Света. Ряд видов этого рода достигает на севере Японии и южных районов Дальнего Востока. Наконец, следует упомянуть палеотропический род колеус. Некоторые виды, в том числе колеус съедобный (С. edulis), имеют крахмалистые клубневидно утолщенные корни и культивируются как пищевые растения в тропиках Старого Света. Многие виды декоративны, и некоторые из них выращивают в комнатах и садах. Индо-малайский вид колеус амбоинский (С. amboinicus) употребляют в качестве приправы к еде, а корни индийского колеуса веттивериовидного (С. vettiverioides) идут на различные украшения. В подсемейство катофериевые (Catopherioideae) входит лишь один род катоферия (Catopheria, 3 вида), распространенный от Мексики до Колумбии. Виды катоферии — очень оригинальные по облику растения, характеризующиеся зародышем с сочным корешком, прилегающим к семядолям. Подсемейство лавандовые (Lavanduloideae) также заключает только один род лаванда (Lavandula). Род лаванда, насчитывающий около 28 видов, распространен главным образом в Средиземноморье и Макаронезии, но его ареал простирается до Сомали в Африке и до Индии. Сюда входят полукустарники и кустарники. Некоторые виды с древних времен используют для получения ценных эфирных масел. Лаванда узколистная (L. angustifolia) — кустарник до 1 м, а иногда до 2 м высотой, широко культивируется для получения ценных эфирных масел и очень популярен также как декоративное растение. Эфирные масла получают также из лаванды широколистной (L. lati-folia) и некоторых других видов. Сухие цветки и листья лаванды долго сохраняют пряный запах, их используют для отпугивания моли. Следующее подсемейство — празиевые (Ргаsioideae) состоит из 6 родов, распространенных преимущественно в тропической Азии. Лишь один монотипный род празиум (Prasium) встречается в Средиземноморье от Португалии до Югославии. Для празиума, как и для других представителей подсемейства, характерны костянковидные доли плода.
К обширному подсемейству яснотковые (Lamioideae, у Мельхиора оно называется Stachyoideae) принадлежит подавляющее большинство цветков. В это подсемейство входят только два рода — большой род шлемник (Scutellaria), насчитывающий около 300 видов, распространенный очень широко по земному шару (за исключением Южной Африки), и монотипный род салазария (Salazaria), распространенный в США и Мексике. Более естественной по сравнению с системой Мельхиора является система губоцветных, предложенная в 1967 г. Р. Вундерлих. Она основывается главным образом на строении плодов и пыльцевых зерен, а в последнее время находит подтверждение и данными хемотаксономии. Вундерлих принимает только 6 подсемейств: простантеровые, живучковые, шлемниковые, чистецовые, чаберовые (Saturejoideae) и катофериевые. Подсемейство празиевых Мельхиора она объединяет с чистецовыми, а подсемейства розмариновые и лавандовые — с выделяемым ею из яснотковых (чистецовых) подсемейством чаберовых. Базиликовые Мельхиора также присоединяются Вундерлих к чаберовым, но занимают в этом подсемействе обособленное положение. Несмотря на то что филогенетическая система Вундерлих имеет ряд достоинств, она также, несомненно, подвергнется дальнейшим изменениям.
Порядок губоцветные (Lamiales) (Н. Н. Цвелев)
Семейство вербеновые (Verbenaceae)
Во внетропических странах семейство вербеновых не пользуется большой известностью. Лишь вербена гибридная (Verbena hybrida), широко культивируемая в качестве декоративного растения, знакома очень многим. Не считая нескольких одичавших и занесенных из других стран видов этого семейства, только 2 вида вербены, витекс священный (Vitex agnuscastus, рис. 213) и фрима тонкокистевая (Phryma leptostachya), произрастают дико на территории нашей страны. Однако вербеновые - довольно большое семейство, по числу видов лишь немного уступающее широко представленному в нашей стране близкому семейству губоцветных и играющее существенную роль в растительности тропических стран.
В противоположность губоцветным среди вербеновых кустарники и деревья значительно преобладают над травами. Некоторые деревья, например тектона большая (Tectona grandis, рис. 212), достигают в высоту более 50 м. Кустарники и невысокие деревья из широко распространенного в тропиках рода авиценния (Avicennia, см. рис. 213) являются важной составной частью мангровых зарослей на затопляемых приливами побережьях морей и океанов. Подобно некоторым другим видам этих замечательных во многих отношениях группировок растительности, они имеют особые, дыхательные корни - пневматофоры, растущие вертикально вверх, выступающие из ила во время отлива и снабжающие подземные части растения кислородом.
Среди других кустарниковых вербеновых имеются как "эрикоидные" (верескоподобные) формы с очень мелкими кожистыми листьями, так и оригинальные безлистные или почти безлистные прутьевидные формы, иногда имеющие ширококрылатые стебли. Особенно интересен внетропический южноамериканский кустарничек неоспартон хвойниковидный (Neosparton ephedroides), имеющий почти безлистные членистые стебли с супротивно или мутовчато расположенными ветвями и внешне чрезвычайно сходный с(голосеменным растением эфедрой.
Многие вербеновые - кустарниковые лианы, обычно лазящие, иногда с помощью придаточных корней-подпорок, реже вьющиеся (например, клеродендрум Томсона - Clerodendrum thomsoniae). У лантаны лиловой (Lantana lilacina) 2 или все 4 ребра четырехгранного стебля сильно разрастаются, вследствие чего стебли становятся лентовидными или четырехкрылыми. Позднее средняя часть стебля разрушается и 2 или 4 части первоначально единого стебля отделяются друг от друга, как бы становясь самостоятельными стеблями.
Все травянистые вербеновые принадлежат к многолетникам. Настоящих однолетников нет совсем, хотя некоторые культивируемые или заносные виды тропического происхождения в условиях умеренно теплого климата ведут себя как однолетники. Преобладают травы с прямостоячими четырехгранными стеблями, но встречаются и другие жизненные формы. Так, у широко распространенного в тропиках и заходящего в качестве заносного растения на юг СССР сорняка липпии ползучей (Lippia repens) почти нитевидные стебли стелются по земле и укореняются в узлах.
Листья вербеновых обычно супротивные, реже очередные или мутовчатые (например, у культивируемой алоизии трехлистной - Aloysia triphylla), цельные или различным образом рассеченные, всегда без прилистников. Для одного из наиболее крупных родов семейства - витекс (Vitex) - характерны пальчатосложные листья с (1)3-7 листочками. Иногда листья сильно видоизменены вплоть до полной их редукции у некоторых прутьевидных кустарников и кустарничков. У вербены спаржелистной (Verbena asparagifolia) листья целиком видоизменены в трехраздельную колючку, а у подушкообразующих вербен из секции пунгентес (Pungentes) тесно скученные на укороченных стеблях кожистые листья оттянуты на верхушке в довольно длинную колючку. У клеродендрума колючего (Clerodendrum aculeatum) колючками становятся черешки листьев после опадания пластинок.
На листьях, как и на других частях растения, обычно имеется опушение из простых, железистых, чешуевидных или звездчатых волосков, но встречаются и совершенно лишенные его виды. У многих клеродендрумов на нижней стороне листьев вдоль средней жилки имеются внецветковые нектарники. Листья американского рода петрея (Petrea) интересны присутствием на них светочувствительных органов, называемых оцеллами. Это особые волоски, выпуклое основание которых находится в глубоких ямках, и только их конусовидная верхушка возвышается над эпидермисом. У видов авиценнии, обитающих в мангровых зарослях, на нижней стороне листьев имеются особые устьица, служащие для выведения избытка солей. В сухую погоду выделенная ими соль покрывает сплошным слоем нижнюю поверхность листьев, придавая им сероватый оттенок. Большинство видов вербеновых имеет диацитные устьица, редко они парацитные или аномоцитные.
Обычно пятичленные цветки вербеновых объединены в различным образом построенные и большей частью имеющие прицветники соцветия верхоцветного или бокоцветного типа. В последнем случае они могут быть кистевидными, колосовидными (например, у многих видов вербены) или головковидными (например, у липпии ползучей). В соцветиях верхоцветного типа полузонтики (дихазии) могут быть расположены в пазухах листьев или в пазухах прицветников, часто образуя метелковидные, щитковидные или зонтиковидные соцветия. Прицветники в соцветиях вербеновых нередко окрашены и принимают участие в привлечении насекомых-опылителей. Так, у некоторых видов лантаны (Lantana) яркоокрашенные прицветники образуют обертку, окутывающую головкообразное соцветие.
Цветки вербеновых более или менее зигоморфные, причем чашечка гораздо чаще сохраняет актиноморфное строение, чем венчик. Совершенно актиноморфные цветки с пяти- или шестичленным околоцветником известны лишь у малезийского рода гёнсия (Geunsia), который имеет также 5-6 далеко выступающих из венчика тычинок. Чашечка всегда сростнолистная и имеет трубчатую колокольчатую или воронковидную форму. Число ее зубцов или лопастей варьирует от 2 до 8, но обычно равно 5. Нередко зубцы слабо развиты, а у палеотропического рода холмшелъдия (Holmskioldia) окраина очень широковоронковидной чашечки совсем лишена зубцов. У индо-малезийского рода гименопирамис (Hymenopyramis) сильно разрастающаяся при плодах чашечка окрашена в белый цвет и имеет 4 широких продольных крыла. Довольно крупные и окрашенные в белый цвет чашечки при ярко-красных венчиках имеет и широко культивируемый в оранжереях африканский клеродендрум Томсона (Glerodendrum thomsoniae). Очень оригинально строение чашечек у индонезийского клеродендрума Минахассы (C. minahassae). He только перед цветением, но и во время него они вздуты, плотно замкнуты и заполнены водой, выделяемой водяными устьицами - гидатодами. Такие чашечки хорошо защищают внутренние части цветка и нектар от нападений насекомых, не являющихся опылителями. Некоторые другие вербеновые для защиты цветков и находящегося в них нектара от непрошеных гостей прибегают к помощи муравьев. У яванской гмелины прицветниковай (Gmelina bracteata) на чашечках имеются внецветковые нектарники, которые очень активно посещаются муравьями. Настоящим растением-муравьелюбом является клеродендрум мирмекофилъный (Glerodendrum myrmecophilum), полые междоузлия стеблей которого служат постоянными жилищами для муравьев. Интересно, что эти жилища имеют специальные входы на небольших выростах в верхней части междоузлий. На каждом таком выросте имеется участок очень тонкой ткани, легко прогрызаемой муравьями при первом заселении ими жилища.
Строение сростнолепестного венчика цветка вербеновых варьирует еще в большей степени, чем строение чашечки: от актиноморфного (у гёнсии) до двугубого и очень сходного с венчиком губоцветных. Очень варьируют и размеры венчика, достигающего у некоторых видов клеродендрума, опыляемых длиннохоботковыми ночными бабочками, длины 15 см. Две нижние лопасти венчика имеют тенденцию к слиянию, и потому многие вербеновые имеют четырехчленный венчик. Некоторые виды клеродендрума имеют мешковидно-разросшуюся нижнюю губу венчика, в других случаях (у бразильского монохилуса - Monochilus) венчик в нижней части, напротив, глубоко расщеплен. Внутри трубки венчика нередко имеются приспособления для сохранения нектара - нек-таростегии. Так, у премны (Premna) внутренняя поверхность трубки покрыта волосками, а у хлоантеса (Chloanthes) в нижней части трубки имеется кольцо из густо расположенных ветвистых волосков. Окраска венчиков может быть очень различной, но преобладают красные, розовые и лиловые цвета. У культивируемой в оранжереях лантаны камара (Lantana camara) венчики меняют свою окраску 3 раза: у только что распустившихся цветков они оранжевые, затем становятся желтыми и к концу цветения - темно-красными.
Число тычинок в цветках вербеновых может быть равно 5 (например, у тектоны), но обычно имеются только 4 тычинки (иногда еще стаминодий пятый), прикрепленные к трубке венчика. У австралийской маллофоры (Mallophora) нити тычинок очень короткие (пыльники почти сидячие) и прикреплены к трубке венчика между лопастями венчика. У тектоны, напротив, более длинные нити тычинок прикреплены почти у основания трубки венчика. Одна из пар тычинок часто менее развита и нередко представлена стаминодиями.
Пыльники вербеновых всегда интрорзные с параллельными или расходящимися в стороны гнездами. У американского рода стахитарфета (Stachytarpheta) гнезда пыльников настолько разошлись, что расположены в одну линию и пыльники производят впечатление одногнездных.
Гинецей вербеновых образован обычно 2, реже 4 (у дюранты - Duranta) или 5 (у гёнсии - Geunsia) плодолистиками. У лантаны, липпии и некоторых других родов задний из двух плодолистиков редуцируется. Образуется псевдомономерный гинецей, который, однако, делится ложной перегородкой пополам с одним семязачатком в каждой части и становится вторично двугнездным. У очень многих родов с двуплодолистиковым гинецеем каждое из двух первичных гнезд также разделяется ложной перегородкой на 2 односемянных гнезда и гинецей становится вторично четырехгнездным. Столбик обычно отходит от верхушки завязи, реже более или менее погружен между ее лопастями, как у близкого семейства губоцветных.
Перекрестному опылению почти всегда обоеполых цветков вербеновых способствует протандрия, а в некоторых случаях и гетеростилия. Только у распространенного в тропической Америке рода эгифила (Aegiphila) имеется неполная двудомность: одни особи несут мужские цветки с далеко выступающими из трубки венчика тычинками и недоразвитым гинецеем; другие - женские цветки с рудиментами тычинок и хорошо развитым гинецеем.
Плодам вербеновых придается большое значение при разделении этого семейства на подсемейства и трибы. Обычный тип плода - костянка с 2-4 косточками и мясистым или сочным эндокарпием. У богатого видами рода вербена и некоторых других родов сухой плод распадается на 4 орешкообразные доли. Еще целый ряд родов имеет плод-коробочку, раскрывающуюся 2 или 4 створками. Чашечка обычно остается при плодах и нередко сильно разрастается. Семена имеют прямой зародыш и обычно лишены эндосперма (он имеется у авиценнии и немногих других родов). Авиценнии свойственна еще одна замечательная особенность семян, связанная с обитанием в специфических условиях мангровых зарослей. Прорастание семян у нее начинается уже на материнском растении и опадающие односемянные (вследствие редукции 3 остальных семязачатков) плоды несут не обычный зародыш, свойственный другим вербеновым, а вполне развитый проросток.
По новейшим данным, вербеновые включают около 100 родов и 3000 видов, распространенных почти исключительно в тропических и субтропических странах обоих полушарий. Лишь немногие роды (вербена, витекс и др.) заходят в южные районы умеренно теплых областей, где они представлены также лишь немногими видами. Наиболее богаты вербеновыми Юго-Восточная Азия с Малайским архипелагом (здесь много эндемичных родов), Центральная Америка с островами Карибского моря и Южная Америка. В тропической Африке разнообразие вербеновых менее велико, однако роды клеродендрум и витекс здесь представлены многочисленными видами. Кроме того, 3 небольших рода эндемичны для Мадагаскара, 1 род - для острова Сокотра, а род стильба (Stilbe) с 4 близкими родами - для Южной Африки.
Среди вербеновых много растений тропических лесов, в том числе лесообразующих пород (виды тектоны, витекса и др.). Виды авиценнии, как уже упоминалось, принадлежат к характернейшим растениям мангровых зарослей почти во всех тропических странах. Кроме того, это единственный род мангровых растений, выходящий за пределы тропиков на территории Новой Зеландии. Невысокий кустарник витекс священный, или "авраамово дерево" (см. рис. 213), играет заметную роль в растительности побережья Средиземного моря. В Южной Америке многие вербеновые являются нагорными ксерофитами, нередко имеющими очень оригинальные жизненные формы (колючие подушки, прутьевидные кустарники и т. д.).
Взаимоотношения между цветками вербеновых и их опылителями разнообразны, но еще недостаточно изучены. Виды родов с короткой трубкой венчика обычно опыляются мелкими перепончатокрылыми и мухами, с более длинной трубкой - крупными перепончатокрылыми и бабочками. Наиболее крупноцветковые виды опыляются длиннохоботковыми бабочками и птицами (в Америке - колибри, а Азии, Африке и Австралии - нектарницами и медососами). Цветки, опыляемые птицами, лишены запаха и обычно имеют наиболее привлекательную для птиц красную окраску венчика. К орнитофильным видам принадлежит разводимый в оранжереях гималайский кустарник холмшелъдия кроваво-красная (Holmskioldia sanguinea). Ее цветки имеют не только красный венчик, но и окрашенную в темно-красный цвет очень широкую чашечку. 4 тычинки и рыльце выступают из трубки венчика и касаются головки парящей возле цветка птицы. В последнее время выяснено, что меняющие свою окраску цветки лантаны камара опыляются очень мелкими насекомыми - пузыреногими или трипсами. Трипсы постоянно встречаются только внутри желтых цветков, которые имеют вполне развитые рыльца. Таким образом, смена окраски цветков этого вида указывает опылителям, в какое время следует посещать цветки.
Виды родов вербеновых, имеющих плоды-костянки, в том числе таких богатых видами, как клеродендрум и витекс, распространяются с помощью птиц. Ярко окрашенные чашечки многих видов клеродендрума служат дополнительным средством для их привлечения. Способы распространения видов с сухими плодами еще мало изучены. Доли плода европейского руде-рального сорняка вербены лекарственной (Verbena officinalis) легко переносятся с почвой на ногах человека и крупных животных, хотя сильно разветвленные сухие стебли этой вербены, по-видимому, могут переноситься и ветром, рассеивая плоды по типу перекати-поле. Плоды обитающей на побережьях морей и океанов авиценнии распространяются водой во время приливов и отливов, а на более далекие расстояния во время штормов.
Значение вербеновых в жизни человека достаточно велико. Прежде всего к этому семейству принадлежат многие тропические деревья, дающие очень ценную древесину. Особенность этой древесины связана с наличием в ней специфических смол. Наиболее известное из них - тектона большая или тековое дерево (см. рис. 212). В некоторых районах Индии, Бирмы, Таиланда и острова Ява она является лесообразующей породой влажных тропических листопадных лесов. В настоящее время тектона культивируется не только в Южной и Юго-Восточной Азии, но также в тропической Африке и Южной Америке. Ее древесина отличается красотой, высокой прочностью и устойчивостью против гниения и повреждения насекомыми, легко поддается обработке. Используется она главным образом в судостроительной и вагоностроительной промышленности, а также для изготовления мебели, составляя важную статью экспорта стран тропической Азии. Очень твердую древесину дают принадлежащие к числу так называемых "железных деревьев" виды американского рода цитарексилум (Citharexilum). Очень высокими качествами обладают и древесины некоторых тропических видов витекса. Средиземноморский вид этого рода витекс священный широко культивируется в качестве декоративного кустарника. Его гибкие ветви используются для плетения различных изделий. Кроме того, он содержит эфирные масла и прежде использовался в народной медицине. Плоды этого вида могут служить заменителем перца, а мясистые костянки некоторых тропических видов, например африканского витекса Момбассы (V. mombassae) довольно широко используются в пищу местным населением.
Ценное эфирное масло, обладающее приятным (напоминающим лимон) запахом и потому используемое в парфюмерии и пищевой промышленности, дает алоизия трехлистная (Aloysia triphylla, см. рис. 212), называемая также "лимонной вербеной". Этот южноамериканский кустарник с мутовчатыми листьями иногда культивируется в садах и парках юга СССР. Многие другие кустарниковые вербеновые принадлежат к числу декоративных растений. Среди них много видов клеродендрума (клеродендрум Томсона, клеродендрум Бунге - C. bungei и др.), часто имеющие ярко окрашенные чашечки. Это наиболее крупный род семейства (более чем с 400 видами). Нередко встречаются в культуре виды лантаны, особенно лантана камара (Lantana camara) и красивоплодника (Callicarpa), имеющие сине-фиолетовые или пурпурно-фиолетовые плоды-костянки. Несколько травянистых декоративных растений имеется в роде вербена (Verbena), включающем более 300 видов. Наиболее известна из них вербена гибридная (V. hybrida), происходящая от гибридизации нескольких южноамериканских видов и культивируемая на значительной территории нашей страны. Другие виды встречаются в культуре значительно реже. Некоторые вербены имеют лекарственное значение. Так, вербена крапиволистная (V. urticifolia) применяется как жаропонижающее и тонизирующее средство. Прежде в народной медицине широко использовалась и европейская вербена лекарственная (V. officinalis).
Семейство губоцветные (Lamiaceae, или Labiatae)
Представители этого семейства легко узнаются уже по строению венчика цветков, имеющего длинную трубку и двугубый зев, напоминающий разинутую пасть сказочного животного. Хотя сходный по облику венчик имеется еще в нескольких семействах цветковых растений (например, у многих норичниковых), для губоцветных он особенно характерен. Не менее своеобразен и плод губоцветных, состоящий из 4 односемянных орешкообразных или редко костянковидыых долей, тогда как у сходных с ними по строению венчика норичниковых плод - обычно многосемянная коробочка. К числу важных отличительных особенностей губоцветных принадлежат также супротивные (очень редко мутовчатые), обычно цельные листья без прилистников и обычно четырехгранные стебли. Очень существен ароматический запах, свойственный большинству видов губоцветных, который определяется присутствием на всех или на некоторых частях растения желёзок, выделяющих эфирные масла сложного состава (в них входят ароматические спирты, фенолы, терпены, альдегиды и другие органические соединения). Именно присутствием этих масел в значительной степени определяется практическое использование губоцветных в качестве технических, лекарственных и ароматических растений.
Большинство губоцветных - травы и полукустарники. Однако среди них, особенно в тропиках и субтропиках, много и кустарников, примером которых может служить распространенный в Средиземноморской флористической области розмарин лекарственный (Rosmarinus officinalis, табл. 55) - вечнозелёный кустарник с мелкими линейными листьями и сине-фиолетовыми (до почти белых) цветками (рис. 214). Губоцветные - деревья имеются только в тропиках, но, в отличие от близкого преимущественно древесного семейства вербеновых, ими являются лишь немногие виды двух американских родов: хиптис (Hyptis) и левкосцептрум (Leucosceptrum). "Чемпион" по высоте среди них - бразильский вид хиптис перепончатый (Н. membranacea), достигающий в высоту 12-15 м, в то время как другие древесные губоцветные обычно не достигают высоты 5 м. В тропиках встречаются и немногие лианы, к которым принадлежат лишь американский род салазария (Salazaria), некоторые виды шлемника (Scutellaria, табл. 55) и гавайского рода стеногина (Stenogyne).


Стебли травянистых губоцветных обычно прямостоячие и не нуждаются в опоре, хотя имеются виды со стелющимися по земле и укореняющимися в узлах стеблями (например, будра плющелистная - Glechoma hederacea). У живучки ползучей (Ajuga reptans), кроме прямостоячих репродуктивных побегов, из пазух листьев розетки образуются дугообразные направленные к земле и укореняющиеся верхушками вегетативные побеги, подобные усам земляники. Хорошо развитая розетка прикорневых листьев, сохраняющаяся во время цветения растения, имеется у многих травянистых губоцветных (например, у некоторых шалфеев - Salvia).
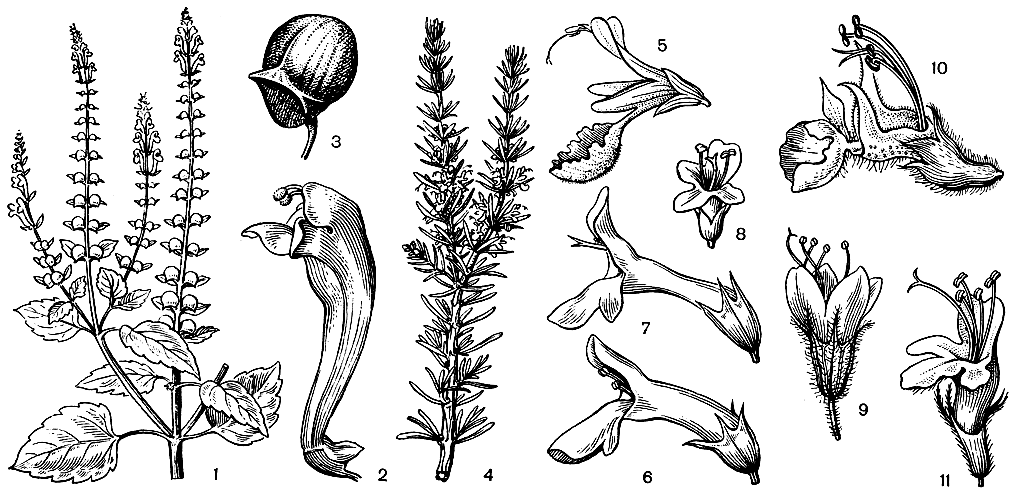
Главный корень часто сохраняется в течение всей жизни растения, реже отмирает и замещается придаточными корнями, отходящими или от основания стебля или от отходящих от него ползучих подземных побегов - корневищ, свойственных многим видам губоцветных. Довольно редко среди губоцветных встречаются виды с корневыми отпрысками, например живучка женевская (Ajuga gennevensis, табл. 55). У многих прибрежных видов, обитающих на переувлажненных местообитаниях, в корневищах образуются воздухоносные полости или участки воздухоносной ткани. Некоторые губоцветные имеют клубневидно утолщенные корни, в тропических странах употребляемые в пищу.

При супротивном расположении листьев близлежащие пары крестообразно чередуются друг с другом. Следствием этого и является четырехгранность стеблей губоцветных, причем грани могут быть плоскими, выпуклыми или вогнутыми. Видов, имеющих мутовчатое расположение листьев, среди губоцветных немного. К ним принадлежит, в частности, оригинальный австралийский кустарник вестрингия (Westringia) с мелкими цельнокрайними листьями, расположенными мутовками по 3-6 (рис. 215). Очередное листорасположение отмечено только у первых листьев проростков в родах зопник (Phlomis) и буквица (Betonica).
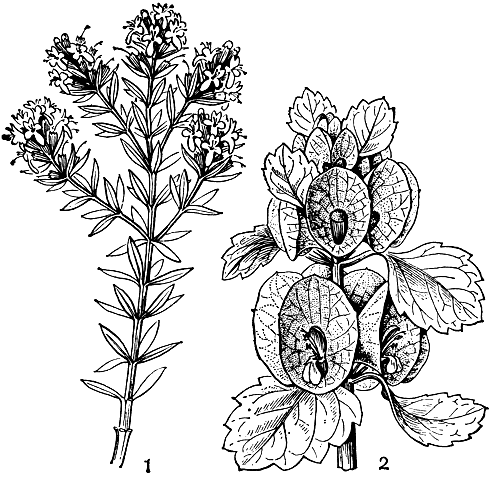
Листья губоцветных обычно цельные и часто цельнокрайние, хотя встречаются и перисто-раздельные (например, у шалфея скабиозолистного - Salvia scabiosifolia). Известны как голые или почти голые виды, например декоративный шалфей блестящий (S. splendens), так и виды, густо покрытые волосками. Среди последних такие средиземноморские виды, как чистец критский (Stachys cretica) и железница крымская (Sideritis taurica), не уступают по красоте знаменитому альпийскому эдельвейсу. Из волосков наиболее распространены многоклеточные простые волоски. Головчатые волоски, головка которых функционирует как желёзка, вырабатывающая эфирное масло, встречаются у многих губоцветных.
Обычно пятичленные и, как правило, обоеполые цветки губоцветных располагаются в пазухах неизмененных или видоизмененных в прицветники листьев. Лишь в немногих случаях (например, у видов шлемника) они одиночные, обычно же цветки образуют так называемые ложные мутовки, составленные двумя супротивными соцветиями, несущими прицветнички. Оси всех первичных соцветий, часто сильно укороченные и составляющие их цветки, кажутся расположенными непосредственно в листовых пазухах, образуя колосовидные соцветия. Иногда ось общего соцветия сильно укорочена и все соцветие становится головкообразным, например у зизифоры головчатой (Ziziphora capitata) и у большого американского рода хиптис. У нередкого в горных районах Средней Азии зайцегуба (Lagochilus) расположенные при основании ложных мутовок прицветнички видоизменены в мощные колючки, защищающие растение от поедания травоядными животными. У некоторых других губоцветных в колючки видоизменяются прицветники или верхние листья, а иногда зубцы листьев.
И чашечка, и венчик губоцветных обычно образованы 5 сросшимися своей основной частью в трубку листочками. Лишь немногие роды, например западносредиземноморский род преслия (Preslia), имеют четырехчленный околоцветник. Чашечка губоцветных может иметь самую разнообразную форму: трубчатую, колокольчатую, ворончатую, шаровидную, а в зеве может быть как двугубой без каких-либо зубцов, так и 5(4)-зубчатой с зубцами одинаковой или различной длины (в последнем случае чашечка также более или менее двугубая). В отличие от венчика, видоизменения которого связаны с приспособлением к опылению, видоизменения чашечки обычно связаны с приспособлением к распространению плодов. Нередко (например, у пустырника) зубцы чашечки имеют вид отогнутых в сторону колючек. Иногда вся чашечка или ее зубцы сильно разрастаются, увеличивая парусность чашечки при распространении семян с помощью ветра, или чашечка становится ярко окрашенной, играя роль в привлечении насекомых или птиц-опылителей, например, ярко-красная чашечка шалфея блестящего. Очень оригинальна чашечка у большого (около 300 видов), почти космополитного рода шлемник. Она здесь двугубая с цельно-крайними губами и после созревания плода распадается на 2 части, имеющие вид створок: нижнюю остающуюся и верхнюю опадающую. Верхняя часть чашечки у многих видов этого рода несет еще поперечную чешуевидную складку - щиток. Шлемник имеет еще ряд особенностей, отличающих этот род от других родов губоцветных (в том числе отсутствие эфиромасличных желёзок), и не случайно некоторые авторы даже предлагали выделить его в особое семейство шлемниковые (Scutellariaceae).
Как уже упоминалось выше, венчик губоцветных обычно разделен на две губы, из которых верхняя образована 2, а нижняя - 3 лепестками. Верхняя губа может быть плоской или выпуклой, иногда она бывает цельнокрайней, так что никаких следов присутствия 2 лепестков не обнаруживается. Нижняя губа почти всегда более крупная (посадочная площадка для опылителей), трехлопастная с более крупной и нередко, в свою очередь, двулопастной средней лопастью. Иногда на ее боковых лопастях имеются нитевидные придатки, как у яснотки (Lamium). Довольно оригинальное строение венчика у родов дубравник (Teucrium) и живучка (Ajuga, табл. 55). У первого из них верхней губы нет совсем и тычинки вместе со столбиком далеко выступают из зева венчика (рис. 214). Однако 2 верхние лопасти, обычно образующие верхнюю губу, здесь не исчезли, а присоединены к нижней губе венчика, составленной не из 3, а из 5 лопастей. У живучки верхняя губа очень короткая по сравнению с длинной нижней губой и венчик также кажется одногубым. У базилика (Ocimum) и близких родов верхняя губа венчика образована не 2, как обычно, а 4 лепестками. Нижняя губа состоит всего из одного плоского или вогнутого лепестка. Для родственного базилику шпороцветника (Plectranthus), заходящего на юг Дальнего Востока, характерно, кроме того, наличие вздутия в нижней части трубки венчика, причем у некоторых видов это вздутие переходит в настоящую шпору. Некоторые роды губоцветных, в том числе и зюзник (Lycopus, рис. 214), имеют короткий и почти актиноморфный венчик с 4-5 лопастями. Окраска венчиков у губоцветных может быть розовой, лиловой, сиреневой, синей, желтой, белой, часто в различных комбинациях.
Тычинок в цветках губоцветных обычно 4, прикрепленных к трубке венчика. У тропического рода колеус (Coleus, табл. 55) и некоторых близких родов тычиночные нити срастаются друг с другом, образуя короткую трубку. Иногда имеется рудимент пятой тычинки, вероятно, исчезнувшей в результате преобразования актиноморфного венчика в зигоморфный в ходе эволюции губоцветных. Пара задних тычинок обычно короче передней пары, но иногда, например у котовника (Nepeta), имеет место обратное соотношение. У мяты (Mentha) с ее почти актиноморфным околоцветником все 4 тычинки почти одинаковой длины. Редукция тычинок в пределах семейства идет еще дальше - до 2 тычинок, причем редуцируются 2 задние тычинки, иногда сохраняясь в виде стаминодиев. Две тычинки характерны, например, для средиземноморского рода розмарин, шалфея и североамериканско-мексиканского рода монарда (Monarda). Ниже места прикрепления тычинок нередко имеется волосистое кольцо - защитное приспособление для нектара.

Пыльники губоцветных имеют различную форму. Гнезда их обычно одинаково развиты, реже одно из них (чаще переднее) развито слабее другого или редуцировано. У многих видов шалфея специализация тычинок зашла наиболее далеко в связи с очень совершенным приспособлением цветков к опылению определенными насекомыми (рис. 216). Каждый из пыльников двух имеющихся здесь тычинок превратился в своеобразное рычажное устройство, на одном конце которого находится вполне развитое верхнее гнездо пыльника, а на другом - обычно ложкообразный рудимент второго (нижнего) гнезда пыльника. Разросшийся в длинную нить связник (часть тычинки между гнездами пыльников) подвижно закреплен на очень короткой тычиночной нити. Полная редукция одного из гнезд пыльников у двух верхних тычинок имеет место еще у шлемника и железницы, но удлинения связника здесь не происходит.
Нектарники губоцветных являются производным основания плодолистиков. Наиболее обычный тип нектарника - это диск с 4 лопастями или зубцами. Каждая лопасть может выделять нектар, но эта способность находится в зависимости от степени развития самих лопастей и их проводящей системы. Насекомые находят нектар под завязью в нижней части венчика, однако при обильном выделении нектара им равномерно заполняется вся нижняя часть трубочки венчика и насекомому достаточно опустить хоботок в трубку, чтобы набрать много нектара. У шлемника сплошной нектароносный диск обычно замещен подковообразным нектарником с 3-5 неравными долями.
В строении гинецея всех губоцветных много общего. Он всегда образован двумя плодолистиками с числом гнезд, соответствующим числу плодолистиков. Однако каждое из гнезд делится ложной перегородкой пополам, вследствие чего завязь становится четырехлопастной, с одним семязачатком в каждой лопасти. Столбик у большинства губоцветных отходит от основания лопастей завязи (гинобазический), но в подсемействах живучковых (Ajugoideae) и простантеровых (Prostantheroideae) он обычно не вполне гинобазический или даже отходит почти от верхушки завязи, как в семействе вербеновых. У шлемника завязь не сидячая, как у остальных губоцветных, а расположена на ножке, образованной сильно суженной нижней частью гинецея.
Хотя цветки губоцветных обычно обоеполые, но во многих родах (например, мята, тимьян - Thymus) наряду с ними встречаются и женские цветки с рудиментарными тычинками, обычно имеющие более мелкий и бледно окрашенный венчик. Значительно реже встречаются мужские цветки с рудиментом гинецея (например, у некоторых видов котовника). Клейстогамные цветки с не выступающим из чашечки и обычно не опадающим венчиком можно видеть у обычного во многих районах СССР однолетнего сорняка яснотки стеблеобъемлющей (Lamium amplexicaule). Эти цветки обычно образуются в неблагоприятных климатических условиях: ранней весной или поздней осенью.
Плод губоцветных состоит из 4 односемянных и большей частью орешкообразных долей, имеющих очень различную форму. При плодах венчик обычно опадает (но остается у клейстогамных цветков и у некоторых родов подсемейства живучковых), а чашечка всегда остается и нередко разрастается (особенно у видов рода молуцелла (Molucella, см. рис. 215) и гименократер (Hymenocrater). Эндосперм в зрелых семенах обычно отсутствует, реже сохраняется, что является примитивной особенностью. Наиболее развит эндосперм у видов австралийского подсемейства простантеровых и у рода тетрахондра (Tetrachondra). Наружная оболочка долей плода нередко несет бугорки, сосочки или волоски,что связано со способом их распространения.
По новейшим данным, семейство губоцветных насчитывает около 200 родов и 3500 видов, распространенных почти по всему земному шару. Особенно многочисленны губоцветные в странах древнесредиземноморской флоры - от Канарских островов до Западных Гималаев, где они часто играют заметную роль в растительных группировках. Почти полностью отсутствуют губоцветные в Арктике и Антарктике. Очень мало губоцветных и в зоне тайги. Довольно богаты губоцветными горные районы тропиков, особенно Центральная и Южная Америка. В Австралии представлены главным образом эндемичные для этого континента роды подсемейства простантеровых (6 родов и около 100 видов). Еще беднее губоцветными Новая Зеландия, где имеются лишь по одному виду шлемника и мяты (оба эндемичных) и один из двух видов очень своеобразного рода тетрахондра (второй вид встречается в Патагонии). Род тетрахондра иногда выделяется в отдельное семейство. Относительно богаты губоцветными Гавайские острова с 2 эндемичными родами преимущественно тропического подсемейства празиевых.
Среди губоцветных преобладают нагорные и равнинные ксерофиты на сухих открытых местообитаниях, однако среди них немало и мезофильных лесных и луговых растений. В тропических дождевых лесах представлены лишь немногие виды. Настоящих водных растений среди губоцветных нет совсем, но имеется несколько родов, многие виды которых обитают по берегам водоемов и на болотах. Таковы, например, очень широко распространенные роды мята, зюзник, шлемник.
Взаимоотношения губоцветных с их опылителями-насекомыми (а в тропической и субтропической Америке также колибри) очень сложны и являются результатом длительной сопряженной эволюции. Виды родов с наиболее просто устроенными цветками, имеющими почти правильный венчик с короткой трубкой и 4 тычинки почти одинаковой длины (например, мята см. рис. 214) обычно опыляются мелкими перепончатокрылыми и мухами, так как нектар в них легко доступен. У большинства других губоцветных с хорошо выраженной двугубостью венчика тычинки и столбик прилегают к верхней губе, а нектар помещается в нижней части довольно длинной трубки. Опылителями таких цветков являются преимущественно перепончатокрылые и бабочки, реже крупные мухи из семейства журчалок. Посетители цветка прикасаются спинкой сначала к рыльцу, а затем к пыльникам и уносят на ней часть пыльцы. У яснотки и некоторых других родов высыпание пыльцы на спинку насекомого облегчается присутствием на пыльниках обращенных вниз волосков, за которые насекомое задевает. Замечательные приспособления типа рычага или шлагбаума, который открывает доступ К нектару лишь после попадания пыльцы на спинку насекомого, имеются у видов зопника и черноголовки (Prunella), однако наибольшего совершенства они достигают у видов шалфея, у которых пыльники двух имеющихся тычинок преобразованы в своеобразные подвижные рычажные устройства (см. выше). Опыляющее насекомое, чтобы достать нектар, просовывает голову в трубку венчика, отодвигает внутрь и кверху ложкообразно расширенный рудимент нижнего гнезда пыльника, а находящееся на другом конце удлиненного связника фертильное гнездо пыльника ударяет по спинке насекомого, высыпая на нее пыльцу (см. рис. 216). У американских губоцветных из родов шалфей, шлемник, монарда и других нередки крупные красные цветки, опыляемые крупными ночными бабочками и колибри. Последние, как и бабочки из семейства бражников, парят возле цветков, высасывая клювом нектар и касаясь головкой расположенных под верхней губой или выступающих из венчика рылец и тычинок.
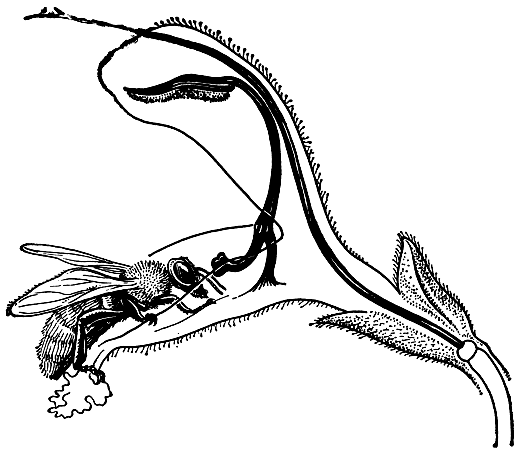
У некоторых губоцветных (особенно у родов подсемейства базиликовых) тычинки и столбик помещаются на нижней губе, так что насекомое, посещающее цветок (обычно бабочки), уносит пыльцу на нижней стороне брюшка. В других родах губоцветных тот же самый эффект (нижнее положение тычинок и столбика) достигается за счет перекручивания трубки венчика (верхняя губа становится как бы нижней губой), перекручивания цветоножки и сильно поникающего соцветия (например, у шалфея поникшего - S. nutans цветущие соцветия обращены вниз верхушкой). Возможность самоопыления цветков губоцветных нередко устраняется за счет более раннего созревания тычинок по сравнению с рыльцем (протандрии), однако во многих других случаях самоопыление вполне возможно. Следует отметить, что у многих губоцветных в привлечении насекомых и птиц принимают участие не только венчик, но и другие части цветка и соцветия. Так, у шалфея блестящего имеются ярко-красные чашечки, а у шалфея дубравного (S. nemorosa) - сине-фиолетовые прицветники.
Очень многие губоцветные распространяются с помощью ветра (анемохория). Единицами распространения - диаспорами - в этом случае обычно являются односемянные части дробного плода, парусность которых может увеличиваться за счет их волосистости или образования крыловидных или хохолковидных выростов. Так, у распространенного в тропической Африке рода тиннея (Tinnea) плоды имеют хохолкообразные щитки. У анемохорных губоцветных стебли часто долго сохраняются в сухом состоянии, постепенно рассеивая плоды (даже в зимнее время). В других случаях, напротив, растопыренно разветвленные стебли с плодоносящими соцветиями легко обламываются у своего основания и перекатываются ветром по степи, постепенно разбрасывая плоды. Такими перекати-поле являются некоторые виды шалфея, зопника, котовника и др. Чем дольше плоды не выпадут из чашечек, тем на большее расстояние они перенесутся. Поэтому у многих губоцветных имеются приспособления для удерживания плодов в чашечке: кольцо волосков в ее зеве или загнутые внутрь зубцы.
У многих анемохорных губоцветных доли плода опадают вместе с чашечкой. Парусность в этих случаях достигается или за счет относительно длинных и нередко реснитчатых зубцов чашечки (например, у тимьяна - Thymus), или за счет сильного разрастания трубки чашечки и ее зубцов. Так, у молуцеллы (Molucella) трубка чашечки при плодах сильно увеличенная, перепончатая, ширококолокольчатая (см. рис. 214), а у зайцегуба, напротив, сильно разрастаются в ширину зубцы чашечки.
У некоторых видов отостегии (Otostegia) роль летучки выполняет сильно расширенная перепончатая верхняя губа чашечки, а у алжирского саккокаликса (Saccocalyx satureioides) чашечка при плодах пузыревидно вздутая с закрытым зубцами зевом, что позволяет находящемуся в ней плоду переноситься ветром на большие расстояния.
Среди губоцветных немало видов, распространяющихся с помощью животных, причем приспособления к зоохории не менее разнообразны. Многие губоцветные имеют ослизняющиеся при смачивании оболочки долей плода и могут распространяться как эндозоохорно (с помощью поедающих плоды животных, преимущественно птиц), так и эпизоохорно (на шерсти и перьях, а также на ногах животных и человека). Большая эффективность эндозоохории достигается путем образования костянковидных долей плода с сочной мясистой оболочкой (у средиземноморского рода празиум). У рода хослундия (Hoslundia) из тропической Америки мясистой (ягодообразной) при плодах становится чашечка, зев которой закрыт зубцами. Эпизоохорно распространяются виды с клейкими или волосистыми долями плода. Во многих случаях для эпизоохорного распространения служат также опадающие вместе с плодами чашечки, жесткие волоски и жесткие оттопыренные в стороны зубцы, которые являются отличным приспособлением для заякоривания в шерсти животных.
Плоды некоторых губоцветных (особенно видов живучки и яснотки) имеют различным образом устроенные придатки, служащие пищей муравьям. Им свойствен так называемый мирмекохорный способ распространения. Бразильский вид хиптис Глазъю (Н. glasiovii) вообще принадлежит к растениям "муравьелюбам" (мирмекофилам): во вздутых междоузлиях его стеблей постоянно находятся поселения особых муравьев.
Виды губоцветных, обитающих у берегов водоемов и на болотах (например, виды мяты и зюзника), имеют плавающие доли плода, приспособленные к распространению водными потоками, но отчасти и водными животными.
Система губоцветных еще далека от совершенства и находится в стадии разработки. Прежде всего еще не вполне ясна граница, отделяющая губоцветных от близкого, но более примитивного семейства вербеновых. Так, некоторые авторы предлагают относить к вербеновым 2 подсемейства губоцветных, сходные по строению гинецея с многими родами вербеновых,- простантеровые и живучковые; другие, напротив, предлагают перенести в губоцветные значительную часть семейства вербеновых. Согласно одной из последних систем семейства губоцветных, разработанной немецким ботаником X. Мельхиором (1964), оно делится на 9 подсемейств. Первое место среди них занимает австралийское подсемейство простантеровые (Prostantheroideae), отличающееся относительно примитивным строением гинецея и семенами с эндоспермом, но имеющее довольно высокоспециализированное строение околоцветника. Далее следует подсемейство живучковых (Ajugoideae), у которого гинецей, как у простантеровых, но семена без эндосперма. Сюда относятся роды живучка, дубравник (Teucrium), аметистея (Amethystea) и др. В особое монотипное подсемейство розмариновые (Rosmarinoideae) выделяется род розмарин (Rosmarinus, табл. 55) с резко выраженным двугубым венчиком, 2 тычинками и семенами без эндосперма (см. рис. 214).
Следующее подсемейство - базиликовые (Ocimoideae), как и все последующие подсемейства, отличается от предыдущих подсемейств более специализированным гинецеем с ясно выраженным гинобазическим столбиком. Тычинок 4, редко 2. Представители этого подсемейства распространены почти исключительно в тропических и субтропических странах. Самый большой род хиптис (Hyptis) заключает более 350 видов, распространенных главным образом в Южной и Центральной Америке. К этому роду относятся самые высокие среди губоцветных деревья, произрастающие в лесах Бразилии. В род хиптис входят два хозяйственно важных вида: хиптис колосоносный (H. spicigera), культивируемый для получения из его семян масла, похожего на кунжутное, и хиптис пахучий (H. suaveolens), или "сангура", дающий очень ароматичный лечебный чай. Род базилик (Ocimum) насчитывает до 150 видов, распространенных в тропических и субтропических странах, особенно в Африке. К этому роду относятся базилик благородный (O. basilicum), родом из тропической Азии, возделываемый во многих странах, в том числе в южных частях СССР, как пряное растение. В Китае культура этого растения известна с 500 г. до н. э. Другой известный представитель этого рода - базилик эвгенольный (O. gratissimum) - кустарник, родом из тропической Азии, культивируется, в частности, у нас в Грузии и в южных районах Краснодарского края как эфиромасличное растение. Пользуется известностью также базилик священный (O. sanctum) - палеотропический кустарник, возделываемый в Индии и в других странах как культурное растение. Род шпороцветник (Pleetranthus) включает около 250 видов, распространенных в тропических и субтропических странах Старого Света. Ряд видов этого рода достигает на севере Японии и южных районов Дальнего Востока. Наконец, следует упомянуть палеотропический род колеус (табл. 55, около 150 видов). Некоторые виды, в том числе колеус съедобный (C. edulis), имеют крахмалистые клубневидно утолщенные корни и культивируются как пищевые растения в тропиках Старого Света. Многие виды декоративны, и некоторые из них выращивают в комнатах и садах. Индо-малайский вид колеус амбоинский (C. amboinicus) употребляют в качестве приправы к еде, а корни индийского колеуса веттивериовидного (C. vettiverioides) идут на различные украшения. В подсемейство катофериевые (Catopherioideae) входит лишь один род катоферия (Catopheria, 3 вида), распространенный от Мексики до Колумбии. Виды катоферии - очень оригинальные по облику растения, характеризующиеся зародышем с сочным корешком, прилегающим к семядолям.
Подсемейство лавандовые (Lavanduloideae) также заключает только один род лаванда (Lavandula). Род лаванда, насчитывающий около 28 видов, распространен главным образом в Средиземноморье и Макаронезии, но его ареал простирается до Сомали в Африке и до Индии. Сюда входят полукустарники и кустарники. Некоторые виды с древних времен используют для получения ценных эфирных масел. Лаванда узколистная (L. angustifolia) - кустарник до 1 м, а иногда до 2 м высотой, широко культивируется для получения ценных эфирных масел и очень популярен также как декоративное растение. Эфирные масла получают также из лаванды широколистной (L. latifolia) и некоторых других видов. Сухие цветки и листья лаванды долго сохраняют пряный запах, их используют для отпугивания моли. Следующее подсемейство - празиевые (Prasioideae) состоит из 6 родов, распространенных преимущественно в тропической Азии. Лишь один монотипный род празиум (Prasium) встречается в Средиземноморье от Португалии до Югославии. Для празиума, как и для других представителей подсемейства, характерны костянковидные доли плода.
К обширному подсемейству яснотковые (Lamioideae, у Мельхиора оно называется Stachyoideae) принадлежит подавляющее большинство внетропических губоцветных. Из представителей этого подсемейства следует прежде всего назвать род погостемон (Pogostemon) с около 40 видами, распространенными в Китае и в тропической Азии. К этому роду относится пачули (P. cablin) - происходящее из Филиппин очень ароматическое растение. Его широко культивируют в тропических странах для получения эфирного масла. Пачулевое масло обладает бактерицидными свойствами и широко используется в парфюмерии и медицине. К числу полезных представителей подсемейства относятся также 5 видов рода перилла (Perilla), распространенных в Южной, Восточной и Юго-Восточной Азии. Перилла кустарниковая (P. frutescens) культивируется в Восточной Азии в качестве масличного и лекарственного растения, а перилла курчавая (P. frutescens var. crispa) с темно-пурпуровыми, курчавыми листьями очень декоративна и в Китае и Японии выращивается как масличная, эфиромасличная и салатная культура. Еще большее значение имеет род мята (Mentha, около 25 видов в умеренной зоне северного полушария, в Южной Африке и в Австралии). Цветки у видов мяты почти актиноморфные, четырехчленные, с 4 почти одинаковыми тычинками. Некоторые виды мяты, особенно гибридная мята перечная (M. piperita), широко культивируют как ценные лекарственные и пищевые (в качестве приправы) растения. Мятные масла, содержащие наряду со многими другими компонентами значительное количество ментола, добавляют во многие лекарственные препараты, в конфеты и в зубную пасту. Иссоп лекарственный (Hyssopus officinalis) также культивируют как эфиромасличное, лекарственное и декоративное растение. Определенное значение имеют также виды рода душица (Origanum). Известно около 15-20 видов этого рода, распространенных в Европе, Средиземноморье и в умеренных областях Азии. Душицу обыкновенную (O. vulgare) используют как лекарственное растение, а листья употребляют как пряность и приправу в пищу и в ликеро-водочном производстве. Широко культивируют майоран (О. majorana), вместе с несколькими близкими видами иногда выделяемый в отдельный род Majorana. Родина майорана - Юго-Западная Азия и Северная Африка. Листья майорана используют в пищу как пряность к различным блюдам и для придания аромата уксусу и чаю. Из листьев и цветков добывают эфирное масло. Одним из наиболее известных представителей семейства является род тимьян (Thymus), насчитывающий от 35 до 400 видов в зависимости от точки зрения систематика на объем вида. Листья тимьяна содержат эфирные масла, главным образом тимол, используемый в медицине. Листья используют как пряность и в консервной и ликеро-водочной промышленности. Средиземноморский тимьян обыкновенный (T. vulgaris) широко культивируется в умеренных и тропических странах. Используются также виды рода мелисса (Melissa, 5 видов в Евразии). Мелиссу лекарственную , или лимонную мяту (M. officinalis), культивируют как эфиромасличное, медоносное и пряное растение. Близок к мелиссе род чабер (Satureja), насчитывающий до 200 видов, распространенных в умеренных и субтропических областях. Чабер садовый (S. hortensis) культивируют как эфиромасличное растение. Его используют как пряность, в медицине и парфюмерии, а также для ароматизации ликеров и коньяков. Наконец, чабер горный (S. montana) разводят как декоративное растение.
Чистец (Stachys) - один из крупных родов подсемейства, насчитывающий около 300 видов, распространенных в умеренных, субтропических и тропических областях, отсутствующих, однако, в Австралии и Новой Зеландии. Некоторые виды чистеца играют заметную роль в сложении растительного покрова (табл. 55). Из числа полезных видов следует упомянуть так называемый китайский артишок (S. affinis), введенный в культуру в Китае и в настоящее время разводимый также в Японии, Бельгии и Франции в качестве овощного растения ради съедобных клубневидных корневищ. Несколько видов чистеца культивируются в качестве декоративных растений.
Шалфей - самый крупный род в семействе губоцветных. Число видов шалфея достигает 700, и они широко распространены в умеренных, субтропических и тропических областях. Некоторые виды шалфея играют заметную роль в растительном покрове. Широко культивируется и всем хорошо известен шалфей лекарственный (S. officinalis). Очень популярным декоративным растением садов и парков стал бразильский шалфей блестящий с ярко-красной чашечкой и венчиком. Мексиканский вид шалфей наркотический (S. divinorum) содержит вещества, обладающие наркотическим действием, что было известно еще древним мексиканцам. В Перу священным цветком считался шалфей супротивноцветковый (S. oppositiflora) - кустарник с красными цветками длиной 2,5-3 см.
Из других лекарственных представителей этого подсемейства укажем также пустырник (Leonurus cardiaca) - известное сердечное средство, кадило (Melittis melissophyllum), виды зизифоры (Ziziphora), зайцегуба.
Последнее место в системе Мельхиора занимает подсемейство шлемниковые (Scutellarioideae), наиболее специализированное по строению цветков. В это подсемейство входят только два рода - большой род шлемник (Scutellaria), насчитывающий около 300 видов, распространенный очень широко по земному шару (за исключением Южной Африки), и монотипный род салазария (Salazaria), распространенный в США и Мексике.
Более естественной по сравнению с системой Мельхиора является система губоцветных, предложенная в 1967 г. Р. Вундерлих. Она основывается главным образом на строении плодов и пыльцевых зерен, а в последнее время находит подтверждение и данными хемотаксономии. Вундерлих принимает только 6 подсемейств: простантеровые, живучковые, шлемни-ковые, чистецовые, чаберовые (Saturejoideae) и катофериевые. Подсемейство празиевых Мельхиора она объединяет с чистецовыми, а подсемейства розмариновые и лавандовые - с выделяемым ею из яснотковых (чистецовых) подсемейством чаберовых. Базиликовые Мельхиора также присоединяются Вундерлих к чаберовым, но занимают в этом подсемействе обособленное положение. Несмотря на то что филогенетическая система Вундерлих имеет ряд достоинств, она также, несомненно, подвергнется дальнейшим изменениям.
Семейство болотниковые (Callitrichaceae)
На поверхности небольших мелководных водоемов нередко можно видеть розетки мелких эллиптических листьев, заканчивающих собой тонкие и гибкие стебли. Это болотник болотный (Callitriche palustris) - наиболее широко распространенный вид единственного рода семейства болотниковых (рис. 217). Болотник нередко называют водяной звездочкой, так как его розетки плавающих листьев действительно напоминают красивую многолучевую звездочку. Как и многие другие виды этого рода, болотник болотный отличается большим полиморфизмом, сильно изменяясь под влиянием различных условий обитания. Наиболее обычная форма растет в воде на глубине до 30-40 см. Стебли его под водой имеют довольно длинные междоузлия и линейные полупрозрачные листья, а у поверхности воды - сближенные междоузлия и значительно более широкие (эллиптические или обратно-яйцевидные) ярко-зеленые плавающие листья, образующие розетку. На более глубоких местах стебли болотника не достигают поверхности воды и несут только подводные листья. В таком виде узнать его нелегко. Но еще более отличается наземная форма болотника болотного. Это карликовое (высотой до 5 см) растеньице с лежачими и укореняющимися в узлах стеблями и очень мелкими, узкоэллиптическими или широколинейными листьями. Наземная форма обычно ведет себя как однолетник, хотя отдельные особи водных форм (особенно более глубоководной) могут существовать по нескольку лет. Представители более высокоспециализированной секции псевдокаллитрихе (Pseudocallitriche) имеют только одну, подводную жизненную форму с широколинейными полупрозрачными листьями, как и у других видов рода, всегда супротивными и не имеющими прилистников.
Однополые, но однодомные цветки болотниковых располагаются по одному или по два в пазухах верхних, а нередко и средних листьев на очень коротких, реже более длинных цветоножках. У их основания часто имеются 2 дуговидно согнутых перепончатых прицветничка. Вследствие перехода к опылению с помощью воды или ветра цветки совершенно лишены околоцветника, так хорошо развитого у близких энтомофильных семейств вербеновых и губоцветных. Каждый женский цветок представлен синкарпным гинецеем из 2 плодолистиков с 2 нитевидными столбиками, переходящими в также нитевидные рыльца. Завязь первично двугнездная, но каждое из двух гнезд делится ложной перегородкой еще на 2 части, из которых каждая содержит по одному висячему анатропному семязачатку. Мужской цветок состоит из одной тычинки. В пазухах листьев могут располагаться 1, реже 2 цветка одного пола, а нередко и разных полов (в последнем случае однополые цветки нетрудно принять за один обоеполый цветок).
Пыльцевые зерна болотниковых шаровидные или эллипсоидальные с 3(4)-бороздной или безапертурной оболочкой. У видов, опыляющихся над поверхностью воды или на ней, пыльцевые зерна имеют тонкую экзину со скульптурой из очень мелких бородавочек, а у видов с подводным опылением экзина совершенно отсутствует. Плоды после созревания распадаются на 4 (реже 2 или 3) односемянные доли.
Хотя род болотник насчитывает всего около 20 видов, он, как и многие другие водные и прибрежные растения, распространен очень широко, являясь почти космополитом (табл. 55). Видов этого рода нет только в высоких широтах Арктики, а также в некоторых пустынях и высокогорьях, хотя отдельные виды поднимаются в горы до 3500 м. Болотники встречаются преимущественно в стоячих и медленно текущих водоемах. Болотник обоеполый (C. hermaphroditica, рис. 217), как и некоторые другие виды секции псевдокаллитрихе, имеющие только подводную форму, выносит слабо солоноватую воду и нередко в изобилии разрастается на песчаном и илистом дне опресненных реками морских заливов на глубине от 0,3 до 2 м. Другие виды также часто встречаются большими группами, заполняя небольшие водоемы.

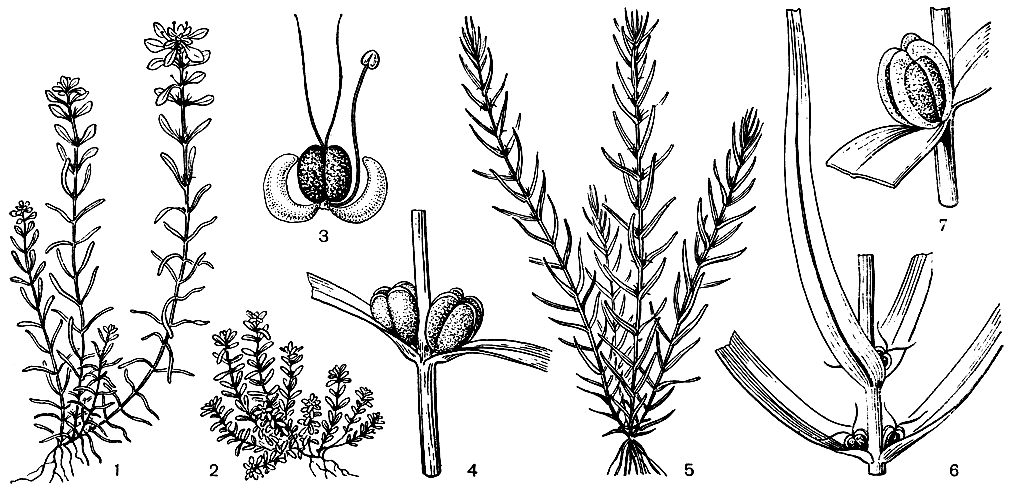
Карликовые наземные формы болотника болотного и многих других земноводных видов этого рода обычно развиваются на месте быстро пересыхающих водоемов, на так называемых вымочках.
У болотников известны 2 способа опыления: надводный и подводный. У большинства видов пыльники на относительно длинных тычиночных нитях и нитевидные рыльца выступают из воды в средней части розеток плавающих листьев. После вскрывания пыльников пыльцевые зерна отчасти падают на поверхность воды, отчасти рассеиваются ветром и в обоих случаях вполне могут попасть на воспринимающую поверхность рылец. Под водой эти виды, как правило, не образуют цветков, хотя созревание плодов у них обычно происходит уже под поверхностью воды.
У видов с подводным опылением, к которым принадлежат облигатно подводные виды секции псевдокаллитрихе, а также водная форма болотника крючковатого (Callitriche hamulata), освобождающиеся из пыльника пыльцевые зерна переносятся потоками воды или длинные нитевидные рыльца непосредственно контактируют со вскрывшимся пыльником. В последнем случае особенно часты случаи опыления женских цветков пыльцой из мужских цветков того же самого побега. При подводном опылении пыльцевые трубки начинают образовываться уже вскоре после выхода пыльцевых зерен в воду, что существенно увеличивает возможность успешных контактов их с рыльцами. Интересно, что у карликовой наземной формы болотника крючковатого опыление также происходит под-водно: или во время временного затопления растений водой, или в каплях дождевой воды, задерживающейся в пазухах листьев. Однако наземная форма более широко распространенного болотника болотного размножается апогамно, образуя большое количество плодов без всякого опыления. Ее цветки обычно имеют лишь рудименты гинецея и недоразвитые пыльники без пыльцевых зерен или с немногими сморщенными зернами.
Доли распадающихся плодов болотника распространяются главным образом постоянными или временными водными потоками. Очень часто доли плодов переносятся еще на ногах животных (в том числе птиц) и человека, о чем можно судить уже по частой встречаемости болотника на пониженных участках дорог и тропинок. У болотника Нафтолъского (C. naftolskyi) и у некоторых других видов с длинными цветоножками отмечена геокарпия: цветоножки после цветения загибаются вниз и сильно удлиняются, погружая созревающие плоды в ил или песок.
По новейшим данным, семейство губоцветных насчитывает около 200 родов и 3500 видов, распространенных почти по всему земному шару. Почти полностью отсутствуют губоцветные в Арктике и Антарктике. Очень мало губоцветных и в зоне тайги. Довольно богаты губоцветными горные районы тропиков.
Среди губоцветных преобладают нагорные и равнинные ксерофиты на сухих открытых местообитаниях, однако среди них немало и мезофильных лесных и луговых растений. В тропических дождевых лесах представлены лишь немногие виды. Настоящих водных растений среди губоцветных нет совсем, но имеется несколько родов, многие виды которых обитают по берегам водоемов и на болотах. Таковы, например, очень широко распространенные роды мята, зюзник, шлемник.
Большинство губоцветных - травы и полукустарники. Однако среди них, особенно в тропиках и субтропиках, много и кустарников. Губоцветные - деревья имеются только в тропиках.
Главный корень часто сохраняется в течение всей жизни растения, реже отмирает и замещается придаточными корнями, отходящими или от основания стебля или от отходящих от него ползучих подземных побегов - корневищ, свойственных многим видам губоцветных. Довольно редко среди губоцветных встречаются виды с корневыми отпрысками. У многих прибрежных видов, обитающих на переувлажненных местообитаниях, в корневищах образуются воздухоносные полости, или участки воздухоносной ткани. Некоторые губоцветные имеют клубневидно утолщенные корни, в тропических странах употребляемые в пищу. При супротивном расположении листьев близлежащие пары крестообразно чередуются друг с другом. Следствием этого и является четырехгранность стеблей губоцветных, причем грани могут быть плоскими, выпуклыми или вогнутыми. Видов, имеющих мутовчатое расположение листьев, среди губоцветных немного.
К числу важных отличительных особенностей губоцветных принадлежат супротивные (очень редко мутовчатые), обычно цельные листья без прилистников и обычно четырехгранные стебли.
Стебли травянистых губоцветных обычно прямостоячие и не нуждаются в опоре, хотя имеются виды со стелющимися по земле и укореняющимися в узлах стеблями (например, будра плющевидная - Glechoma hederacea). У живучки ползучей (Ajuga reptans), кроме прямостоячих репродуктивных побегов, из пазух листьев розетки образуются дугообразные направленные к земле и укореняющиеся верхушками вегетативные побеги, подобные усам земляники. Хорошо развитая розетка прикорневых листьев, сохраняющаяся во время плетения растения, имеется у многих травянистых губоцветных.
Листья губоцветных обычно цельные и часто цельнокрайние, хотя встречаются и перистораздельные. Известны как голые или почти голые виды, так и виды, густо покрытые волосками. Из волосков наиболее распространены многоклеточные простые волоски. Головчатые волоски, головка которых функционирует как желёзка, вырабатывающая эфирное масло, встречаются у многих губоцветных.
Представители этого семейства легко узнаются уже по строению венчика цветков, имеющего длинную трубку и двугубый зев, напоминающий разинутую пасть сказочного животного. Хотя сходный по облику венчик имеется еще в нескольких семействах цветковых растений (например, у многих норичниковых), для губоцветных он особенно характерен.Обычно 5-членные и, как правило, обоеполые цветки губоцветных располагаются в пазухах неизмененных или видоизмененных в прицветники листьев. Лишь в немногих случаях (например, у шлемника) они одиночные, обычно же цветки образуют так называемые.ложные мутовки, составленные двумя супротивными соцветиями, несущими прицветнички. Оси всех первичных соцветий, часто сильно укороченные и составляющие их цветки, кажутся расположенными непосредственно в листовых пазухах, образуя колосовидные соцветия. Иногда ось общего соцветия сильно укорочена и все соцветие становится головкообразным.
И чашечка, и венчик губоцветных обычно образованы 5 сросшимися своей основной частью в трубку листочками. Лишь немногие роды имеют четырехчленный околоцветник. Чашечка губоцветных может иметь самую разнообразную форму: трубчатую, колокольчатую, ворончатую, шаровидную, а в зеве может быть как двугубой без каких-либо зубцов, так и 5 (4) зубчатой с зубцами одинаковой или различной длины (в последнем случае чашечка также более или менее двугубая). В отличие от венчика, видоизменения которого связаны с приспособлением к опылению, видоизменения чашечки обычно связаны с приспособлением к распространению плодов. Нередко зубцы чашечки имеют вид отогнутых в сторону колючек. Иногда вся чашечка или ее зубцы сильно разрастаются, увеличивая парусность чашечки при распространении семян с помощью ветра, или чашечка становится ярко окрашенной, играя роль в привлечении насекомых или птиц-опылителей. Она здесь двугубая с цельнокрайними губами и после созревания плода распадается на 2 части, имеющие вид створок: нижнюю остающуюся и верхнюю опадающую. Верхняя часть чашечки у многих видов этого рода несет еще поперечную чешуевидную складку - щиток.
Как уже упоминалось выше, венчик губоцветных обычно разделен на две губы, из которых верхняя образована 2, а нижняя - 3 лепестками. Верхняя губа может быть плоской или выпуклой, иногда она бывает цельнокрайней, так что никаких следов присутствия 2 лепестков не обнаруживается. Нижняя губа почти всегда более крупная (посадочная площадка для опылителей), трехлопастная с более крупной и нередко, в свою очередь, двулопастной средней лопастью. Иногда на ее боковых лопастях имеются нитевидные придатки, как у яснотки (Lamium). У живучки верхняя губа очень короткая по сравнению с длинной нижней губой и венчик также кажется одногубым. Тычинок в цветках губоцветных обычно 4, прикрепленных к трубке венчика. Иногда имеется рудимент пятой тычинки, вероятно, исчезнувшей в результате преобразования актиноморфного венчика в зигоморфный в ходе эволюции губоцветных. У мяты (Mentha) с ее почти актиноморфным околоцветником все 4 тычинки почти одинаковой длины. Редукция тычинок в пределах семейства идет еще дальше - до 2 тычинок, причем редуцируются 2 задние тычинки, иногда сохраняясь в виде стаминодиев.
Пыльники губоцветных имеют различную форму. Гнезда их обычно одинаково развиты, реже одно из них (чаще переднее) развито слабее другого или редуцировано. Каждый из пыльников двух имеющихся здесь тычинок превратился в своеобразное рычажное устройство, на одном конце которого находится вполне развитое верхнее гнездо пыльника, а на другом - обычно ложкообразный рудимент второго (нижнего) гнезда пыльника. Разросшийся в длинную нить связник (часть тычинки между гнездами пыльников) подвижно закреплен на очень короткой тычиночной нити.
Нектарники губоцветных являются производным основания плодолистиков. Наиболее обычный тип нектарника - это диск с 4 лопастями или зубцами. Каждая лопасть может выделять нектар, но эта способность находится в зависимости от степени развития самих лопастей и их проводящей системы. Насекомые находят нектар под завязью в нижней части венчика, однако при обильном выделении нектара им равномерно заполняется вся нижняя часть трубочки венчика и насекомому достаточно опустить хоботок в трубку, чтобы набрать много нектара.
В строении гинецея всех губоцветных много общего. Он всегда образован двумя плодолистиками с числом гнезд, соответствующим числу плодолистиков. Однако каждое из гнезд делится ложной перегородкой пополам, вследствие чего завязь становится четырехлопастной, с одним семязачатком в каждой лопасти. Столбик у большинства губоцветных отходит от основания лопастей завязи (гинобазический). Хотя цветки губоцветных обычно обоеполые, но во многих родах наряду с ними встречаются и женские цветки с рудиментарными тычинками, обычно имеющие более мелкий и бледно окрашенный венчик. Значительно реже встречаются мужские цветки с рудиментом гинецея.
Взаимоотношения губоцветных с их опылителями-насекомыми очень сложны и являются результатом длительной сопряженной эволюции: виды родов с наиболее просто устроенными цветками, имеющими почти правильный венчик с короткой трубкой и 4 тычинки почти одинаковой длины обычно опыляются мелкими перепончатокрылыми и мухами, так как нектар в них легко доступен. У большинства других губоцветных с хорошо выраженной двугубостью венчика тычинки и столбик прилегают к верхней губе, а нектар помещается в нижней части довольно длинной трубки. Опылителями таких цветков являются преимущественно перепончатокрылые и бабочки, реже крупные мухи из семейства журчалок. Посетители цветка прикасаются спинкой сначала к рыльцу, а затем к пыльникам и уносят на ней часть пыльцы. У яснотки и некоторых других родов высыпание пыльцы на спинку насекомого облегчается присутствием на пыльниках обращенных вниз волосков, за которые насекомое задевает. Замечательные приспособления типа рычага или шлагбаума, который открывает доступ к нектару лишь после попадания пыльцы на спинку насекомого, имеются у видов зопника и черноголовки (Prunella), однако наибольшего совершенства они достигают у видов шалфея, у которых пыльники двух имеющихся тычинок преобразованы в своеобразные подвижные рычажные устройства. Опыляющее насекомое, чтобы достать нектар, просовывает голову в трубку венчика, отодвигает внутрь и кверху ложкообразно расширенный рудимент нижнего гнезда пыльника, а находящееся на другом конце удлиненного связника фертильное гнездо пыльника ударяет по спинке насекомого, высыпая на нее пыльцу.
У некоторых губоцветных тычинки и столбик помещаются на нижней губе, так что насекомое, посещающее цветок (обычно бабочки), уносит пыльцу на нижней стороне брюшка. В других родах губоцветных тот же самый эффект (нижнее положение тычинок и столбика) достигается за счет перекручивания трубки венчика (верхняя губа становится как бы нижней губой), перекручивания цветоножки и сильно поникающего соцветия. Возможность самоопыления цветков губоцветных нередко устраняется за счет более раннего созревания тычинок по сравнению с рыльцем (протандрии), однако во многих других случаях самоопыление вполне возможно. Следует отметить, что у многих губоцветных в привлечении насекомых и птиц принимают участие не только венчик, но и другие части цветка и соцветия.
Не менее своеобразен и плод губоцветных, состоящий из 4 односемянных орешкообразных или редко костянковидных долей, тогда как у сходных с ними по строению венчика норичниковых плод - обычно многосемянная коробочка. При плодах венчик обычно опадает, а чашечка всегда остаётся и нередко разрастается. Эндосперм в зрелых семенах обычно отсутствует, реже сохраняется, что является примитивной особенностью. Наружная оболочка долей плода нередко несет бугорки, сосочки или волоски, что связано со способом их распространения.
Очень многие губоцветные распространяются с помощью ветра (анемохория). Единицами распространения - диаспорами - в этом случае обычно являются односемянные части дробного плода, парусность которых может увеличиваться за счет их волосистости или образования крыловидных или хохолковидных выростов. У анемохорных губоцветных стебли часто долго сохраняются в сухом состоянии, постепенно рассеивая плоды (даже в зимнее время). В других случаях, напротив, растопыренно разветвленные стебли с плодоносящими соцветиями легко обламываются у своего основания и перекатываются ветром по степи, постепенно разбрасывая плоды. Чем дольше плоды не выпадут из чашечек, тем на большее расстояние они перенесутся. Поэтому у многих губоцветных имеются приспособления для удерживания плодов в чашечке: кольцо волосков в ее зеве или загнутые внутрь зубцы.
У многих анемохорных губоцветных доли плода опадают вместе с чашечкой. Парусность в этих случаях достигается или за счет относительно длинных и нередко реснитчатых зубцов чашечки или за счет сильного разрастания трубки чашечки и ее зубцов.
Среди губоцветных немало видов, распространяющихся с помощью животных. Причем приспособления к зоохории не менее разнообразны. Многие губоцветные имеют ослизняющиеся при смачивании оболочки долей плода и могут распространяться как эндозоохорно (с помощью поедающих плоды животных, преимущественно птиц), так и эпизоохорно (на шерсти и перьях, а также на ногах животных и человека). Большая эффективность эндозоохории достигается путем образования костянковидных долей плода с сочной мясистой оболочкой. Эпизоохорно распространяются виды с клейкими или волосистыми долями плода. Во многих случаях для эпизоохорного распространения служат также опадающие вместе с плодами чашечки, жесткие волоски и жесткие оттопыренные в стороны зубцы, которые являются отличным приспособлением для заякоривания в шерсти животных.
Плоды некоторых губоцветных (особенно видов живучки и яснотки) имеют различным образом устроенные придатки, служащие пищей муравьям. Им свойствен так называемый мирмекохорный способ распространения.
Виды губоцветных, обитающих у берегов водоемов и на болотах (например, виды мяты и зюзника), имеют плавающие доли плода, приспособленные к распространению водными потоками, но отчасти и водными животными.
Система губоцветных еще далека от совершенства и находится в стадии разработки. Согласно одной из последних систем семейства губоцветных, разработанной немецким ботаником X. Мельхиором (1964), оно делится на 9 подсемейств.
Очень существен ароматический запах, свойственный большинству видов губоцветных, который определяется присутствием на всех или на некоторых частях растения желёзок, выделяющих эфирные масла сложного состава (в них входят ароматические спирты, фенолы, терпены, альдегиды и другие органические соединения). Именно присутствием этих масел в значительной степени определяется практическое использование губоцветных в качестве технических, лекарственных и ароматических растений.